ธรรมชาติและที่มาของไวรัส อันตรายและประโยชน์ของไวรัส
ไวรัสมีบทบาทสำคัญในชีวิตมนุษย์ เนื่องจากสามารถทำให้เกิดโรคที่มีความรุนแรงต่างกันได้
ตามลักษณะทางระบาดวิทยา โรคไวรัสแบ่งออกเป็น มานุษยวิทยาแล้วมีหลายสิ่งที่ส่งผลกระทบต่อมนุษย์เท่านั้น (เช่นโปลิโอ) และ มนุษย์สัตว์ -ที่ติดต่อจากสัตว์สู่คน (เช่น โรคพิษสุนัขบ้า)
เส้นทางหลักในการแพร่เชื้อไวรัสคือ:
1.เส้นทางอาหารซึ่งไวรัสเข้าสู่ร่างกายมนุษย์ด้วยอาหารและน้ำที่ปนเปื้อน (ไวรัสตับอักเสบ A, E ฯลฯ )
2.ทางหลอดเลือด (หรือทางเลือด) ซึ่ง ไวรัสเข้าสู่เลือดหรือสภาพแวดล้อมภายในของบุคคลโดยตรง สิ่งนี้ส่วนใหญ่เกิดขึ้นเมื่อใช้เครื่องมือผ่าตัดหรือกระบอกฉีดยาที่ปนเปื้อน ในระหว่างการมีเพศสัมพันธ์ที่ไม่มีการป้องกัน และจากแม่สู่ลูกด้วย ไวรัสที่เปราะบางซึ่งสลายตัวอย่างรวดเร็วในสิ่งแวดล้อม (ไวรัสตับอักเสบบี, เอชไอวี, ไวรัสโรคพิษสุนัขบ้า ฯลฯ ) จะถูกส่งผ่านในลักษณะนี้
3.ระบบทางเดินหายใจซึ่งมีลักษณะพิเศษคือกลไกการแพร่เชื้อทางอากาศ โดยที่ไวรัสเข้าสู่ร่างกายมนุษย์พร้อมกับอากาศที่หายใจเข้าไปซึ่งมีอนุภาคของเสมหะและเมือกที่ถูกโยนออกมาโดยคนป่วยหรือสัตว์ นี่เป็นเส้นทางการแพร่เชื้อที่อันตรายที่สุด เนื่องจากไวรัสสามารถแพร่กระจายทางอากาศได้ในระยะทางไกลและทำให้เกิดโรคระบาดทั้งหมด นี่คือวิธีการแพร่เชื้อไวรัสไข้หวัดใหญ่ ไข้หวัดนก คางทูม อีสุกอีใส ฯลฯ
ไวรัสส่วนใหญ่มีความสัมพันธ์บางอย่างกับอวัยวะใดอวัยวะหนึ่ง ตัวอย่างเช่น ไวรัสตับอักเสบจะขยายตัวในเซลล์ตับเป็นหลัก ตามประเภทของอวัยวะเป้าหมายที่ได้รับผลกระทบในระหว่างเกิดโรคใด ๆ เราแยกแยะประเภทของโรคไวรัสดังต่อไปนี้: ลำไส้, ระบบทางเดินหายใจ (ทางเดินหายใจ), ส่งผลกระทบต่อระบบประสาทส่วนกลางและอุปกรณ์ต่อพ่วง, อวัยวะภายใน, ผิวหนังและเยื่อเมือก, หลอดเลือด, ระบบภูมิคุ้มกัน ฯลฯ ตามประเภทการพัฒนาทางคลินิกเราแยกแยะได้ การติดเชื้อไวรัสเฉียบพลันและเรื้อรังที่พบมากที่สุด โรคไวรัสเฉียบพลันที่เกิดขึ้นกับอาการเด่นชัดของท้องถิ่น (ความเสียหายต่อเยื่อเมือกของระบบทางเดินหายใจ, เนื้อเยื่อตับ, ความเสียหายต่อส่วนต่าง ๆ ของสมอง) และลักษณะทั่วไป - อุณหภูมิร่างกายเพิ่มขึ้น, อ่อนแรง, ปวดข้อและกล้ามเนื้อ, การเปลี่ยนแปลง ในองค์ประกอบเลือด เป็นต้น การติดเชื้อไวรัสเฉียบพลันเช่นมักจบลงด้วยการฟื้นตัวของร่างกายอย่างสมบูรณ์ ในบางกรณีรูปแบบเฉียบพลันของโรคจะกลายเป็นเรื้อรัง เรื้อรังการติดเชื้อไวรัสเกิดขึ้นพร้อมกับภาพทางคลินิกที่ไม่ชัดเจนและบางครั้งผู้ป่วยอาจไม่สังเกตเห็นเอง การติดเชื้อเรื้อรังนั้นรักษาได้ยากและอาจคงอยู่เป็นเวลานาน นำไปสู่การเปลี่ยนแปลงทางสัณฐานวิทยาและการทำงานของอวัยวะภายในอย่างมีนัยสำคัญ (เช่น โรคตับอักเสบบีเรื้อรังอาจทำให้เกิดโรคตับแข็งในตับ)
แน่นอนว่าไวรัสรูปแบบใหม่ไม่สามารถเปรียบเทียบกับรูปแบบปฐมภูมิของชีวิตได้ พวกเขาจะต้องแตกต่างจากอย่างหลังอันเป็นผลมาจากการพัฒนาทางประวัติศาสตร์ที่พวกเขาได้รับ แต่ยังคงรักษาคุณสมบัติทั่วไปทั่วไปขององค์กรที่ไม่ใช่มือถือ
ด้วยการค้นพบไวรัสในปี พ.ศ. 2435 โดย D.I. Ivanovsky ช่องว่างในประวัติศาสตร์ของการพัฒนาสิ่งมีชีวิตก็ถูกเติมเต็ม จุดยืนของเองเกลส์เกี่ยวกับพัฒนาการทางประวัติศาสตร์ที่ยาวนานของสิ่งมีชีวิตที่ไม่ใช่เซลล์ได้ถูกทำให้เป็นรูปธรรมในโลกของไวรัส
ลักษณะของไวรัสไวรัสมีตำแหน่งพิเศษเมื่อเปรียบเทียบกับจุลินทรีย์กลุ่มอื่น อย่างที่เคยเป็นมา พวกเขาอยู่บนพรมแดนระหว่างสิ่งมีชีวิตกับธรรมชาติที่ไม่มีชีวิต ธรรมชาติของพวกมันเป็นหัวข้อของการศึกษาและการอภิปรายมากมายตลอดประวัติศาสตร์ของไวรัสวิทยา อะไรเป็นสิ่งมีชีวิตหรือสาร สิ่งมีชีวิตที่มีคุณสมบัติเป็นสารหรือในทางกลับกัน? การตั้งคำถามเหล่านี้บ่งบอกถึงลักษณะที่เป็นสองเท่าของคุณสมบัติของพวกเขา แท้จริงแล้ว ในสภาพแวดล้อมภายนอก ไวรัส รวมถึงฟาจ เป็นรูปแบบที่เฉื่อยโดยสิ้นเชิงซึ่งไม่แสดงสัญญาณของชีวิต เช่น การหายใจ การกินอาหาร และการสืบพันธุ์ หลายแห่งสามารถกลายเป็นคริสตัลได้ ในสถานะผลึกหรือแห้งสามารถเก็บรักษาไว้ได้นานหลายปี พวกมันมีพฤติกรรมเหมือนสารประกอบทางเคมี แต่ทันทีที่พวกเขาเข้าไปในเซลล์ที่บอบบาง สัญญาณของชีวิตทั้งหมดก็เริ่มปรากฏขึ้นในเซลล์เหล่านั้น - การสืบพันธุ์ พันธุกรรม ความแปรปรวน ความสามารถในการปรับตัว วิวัฒนาการ
นักไวรัสวิทยาชาวอเมริกันผู้มีชื่อเสียง ดับเบิลยู. เอ็ม. สแตนลีย์ ถือว่าลักษณะความเป็นคู่ของพวกมันเป็นคุณลักษณะเฉพาะของไวรัส ในสถานะอิสระ ไวรัสเป็นเพียงโมเลกุลขนาดยักษ์ที่มีคุณสมบัติทั้งหมดที่มีอยู่ในโมเลกุลขนาดใหญ่ทั้งหมด ในเซลล์ที่มีชีวิต มันปรากฏตัวเป็นสิ่งมีชีวิต สืบพันธุ์และกลายพันธุ์ ดังนั้น TMV จึงเป็นสิ่งมีชีวิตที่เล็กที่สุดหรือเป็นโมเลกุลที่ใหญ่ที่สุด
นักวิทยาศาสตร์บางคนขึ้นอยู่กับความสามารถของไวรัสในการตกผลึกและเอนไซม์ในการเร่งปฏิกิริยาอัตโนมัติ เช่น การเร่งปฏิกิริยาภายใต้อิทธิพลของผลิตภัณฑ์ที่ปล่อยออกมาอันเป็นผลมาจากปฏิกิริยานั้นเองได้จำแนกไวรัสว่าเป็นเอนไซม์ แต่องค์ประกอบทางเคมีของไวรัสและเอนไซม์จะปฏิเสธธรรมชาติของเอนไซม์ เอนไซม์เป็นสารโปรตีน และไวรัส นอกเหนือจากโปรตีนแล้ว ยังมีกรดนิวคลีอิกที่ควบคุมการสังเคราะห์โปรตีนอีกด้วย ปรากฏการณ์การตกผลึกขึ้นอยู่กับมวลและโครงสร้างของอนุภาคที่ก่อตัวเป็นโครงตาข่ายคริสตัล มวลของแบคทีเรีย ริกเก็ตเซีย และไวรัสขนาดใหญ่นั้นใหญ่เกินกว่าที่พวกมันจะกลายเป็นหน่วยโครงสร้างของผลึก ไวรัสขนาดเล็กที่มีขนาดเล็กสามารถตกผลึกได้เนื่องจากการกระทำของแรงยึดเกาะระหว่างโมเลกุลและโครงสร้างทางเคมีฟิสิกส์ของอนุภาค
การถกเถียงเกี่ยวกับธรรมชาติของไวรัสไม่เพียงแต่เป็นที่สนใจทางทฤษฎีเท่านั้น แต่ยังมีความสำคัญในทางปฏิบัติอีกด้วย เป็นที่ชัดเจนว่ามาตรการในการต่อสู้กับโรคไวรัสของมนุษย์ สัตว์ และพืชจะแตกต่างกันโดยสิ้นเชิง ขึ้นอยู่กับว่าเกิดจากสิ่งมีชีวิตหรือไม่ หรือเกิดขึ้นจากการเข้าสู่ร่างกายหรือการก่อตัวของสารบางอย่างในนั้น . มาตรการทั้งหมดในการต่อสู้กับการติดเชื้อไวรัสนั้นขึ้นอยู่กับการรับรู้ถึงธรรมชาติที่มีชีวิตของไวรัส และการปฏิบัติได้พิสูจน์ประสิทธิผลของมาตรการเหล่านี้แล้ว
แนวคิดสมัยใหม่เกี่ยวกับไวรัสได้รับการพัฒนาอย่างค่อยเป็นค่อยไป ในปี พ.ศ. 2435 ดิ. Ivanovsky ดึงความสนใจไปที่โรคยาสูบที่แพร่หลายซึ่งใบถูกปกคลุมไปด้วยจุดกระจัดกระจาย (โรคโมเสก) หลังจากการค้นพบไวรัสโดย Ivanovsky พวกมันถูกพิจารณาว่าเป็นเพียงจุลินทรีย์ขนาดเล็กมาก ซึ่งไม่สามารถเติบโตบนอาหารเทียมได้ ไม่นานหลังจากการค้นพบไวรัสโมเสกยาสูบ ธรรมชาติของไวรัสของโรคปากและเท้าเปื่อยก็ได้รับการพิสูจน์แล้ว และไม่กี่ปีต่อมาก็มีการค้นพบแบคทีริโอฟาจ ดังนั้นจึงค้นพบไวรัสสามกลุ่มหลักซึ่งแพร่เชื้อให้กับพืช สัตว์ และแบคทีเรีย อย่างไรก็ตาม เป็นเวลานานแล้วที่สาขาไวรัสวิทยาอิสระเหล่านี้พัฒนาขึ้นโดยแยกจากกัน และไวรัสที่ซับซ้อนที่สุด - แบคเทอริโอฟาจ - ได้รับการพิจารณามานานแล้วว่าไม่ใช่สิ่งมีชีวิต แต่เป็นสิ่งที่คล้ายกับเอนไซม์ อย่างไรก็ตามในช่วงปลายทศวรรษที่ 20 และต้นทศวรรษที่ 30 เป็นที่ชัดเจนว่าไวรัสเป็นสิ่งมีชีวิตและในเวลาเดียวกันก็มีการกำหนดชื่อของไวรัสที่กรองได้หรืออัลตราไวรัส
ในช่วงปลายทศวรรษที่ 30 และต้นทศวรรษที่ 40 การศึกษาเกี่ยวกับไวรัสก้าวหน้าไปมากจนความสงสัยเกี่ยวกับธรรมชาติของพวกมันหายไป และแนวคิดเรื่องไวรัสในฐานะสิ่งมีชีวิตก็ถูกสร้างขึ้น พื้นฐานสำหรับการรับรู้ไวรัสว่าเป็นสิ่งมีชีวิตคือข้อเท็จจริงที่ได้รับในระหว่างการศึกษา ซึ่งระบุว่าไวรัส เช่นเดียวกับสิ่งมีชีวิตอื่นๆ (สัตว์ พืช โปรโตซัว เชื้อรา แบคทีเรีย) มีความสามารถในการสืบพันธุ์ มีการถ่ายทอดทางพันธุกรรมและความแปรปรวน มีความสามารถในการปรับตัวเข้ากับ สภาพแวดล้อมที่เปลี่ยนแปลงไป และในที่สุด ความอ่อนไหวต่อวิวัฒนาการทางชีววิทยา ซึ่งเกิดจากการคัดเลือกโดยธรรมชาติหรือโดยธรรมชาติ ก่อนอื่นนี่คือปฏิสัมพันธ์ของจีโนมทั้งสอง - ไวรัสและเซลล์
ตามข้อที่สามไวรัสเป็นอนุพันธ์ของโครงสร้างทางพันธุกรรมของเซลล์ที่ค่อนข้างเป็นอิสระ แต่ยังคงพึ่งพาเซลล์อยู่ สมมติฐานที่สามในช่วง 20-30 ปีดูไม่น่าเป็นไปได้และยังได้รับชื่อที่น่าขันเกี่ยวกับสมมติฐานของยีนบ้าอีกด้วย อย่างไรก็ตาม ข้อเท็จจริงที่สะสมมาทำให้เกิดข้อโต้แย้งใหม่ๆ มากขึ้นเรื่อยๆ ที่สนับสนุนสมมติฐานนี้ นอกจากนี้ ยังมีข้อเท็จจริงจำนวนมากที่สะสมซึ่งบ่งชี้ถึงการมีอยู่ของการแลกเปลี่ยนข้อมูลทางพันธุกรรมจำนวนมาก รวมถึงในหมู่ตัวแทนของไวรัสที่อยู่ห่างไกลจากวิวัฒนาการที่แตกต่างกัน จากการแลกเปลี่ยนดังกล่าว คุณสมบัติทางพันธุกรรมสามารถเปลี่ยนแปลงได้อย่างรวดเร็วและทันทีผ่านการบูรณาการของยีนต่างประเทศ (ยืมฟังก์ชันของยีน) คุณสมบัติทางพันธุกรรมใหม่สามารถเกิดขึ้นได้เนื่องจากการผสมผสานที่ไม่คาดคิดของยีนของตัวเองและยีนที่บูรณาการ (การเกิดขึ้นของฟังก์ชันใหม่) ในที่สุดการเพิ่มขึ้นอย่างง่ายในจีโนมเนื่องจากยีนที่ไม่ทำหน้าที่จะเปิดโอกาสของการวิวัฒนาการของยีนหลัง (การก่อตัวของยีนใหม่)
รูปแบบชีวิตที่ไม่ใช่เซลล์คืออะไร?
มันกัดอย่างเจ็บปวดและน่ารังเกียจ
ถึงแม้บางครั้งจะมองไม่เห็นก็ตาม...
เจ. สวิฟท์
“เอาล่ะ ให้คนแปลกหน้าแสนสวยของเรายังคงเป็นคนแปลกหน้า ตราบเท่าที่เธอรักเรา”? ตามตำนานกล่าวว่านักจุลชีววิทยาที่โดดเด่นแอล. ปาสเตอร์ล้มเหลวในการแยกสาเหตุที่ทำให้เกิดโรคพิษสุนัขบ้า? โรคร้ายแรงซึ่งไม่มีทางรอดในศตวรรษที่ 19 เขาได้รับวัคซีนจึงเข้าใจธรรมชาติของเชื้อโรคและช่วยชีวิตมนุษย์ได้หลายพันคน ในสมัยนั้นไม่มีใครทำเช่นนี้ได้ เนื่องจากสาเหตุของโรคพิษสุนัขบ้าไม่ใช่จุลินทรีย์อย่างที่แอล. ปาสเตอร์คาดหวัง แต่เป็นไวรัส
นอกจากสิ่งมีชีวิตเซลล์เดียวและหลายเซลล์แล้ว ยังมีสิ่งมีชีวิตรูปแบบอื่นในธรรมชาติอีกด้วย เหล่านี้เป็นไวรัสที่ไม่มีโครงสร้างเซลล์ พวกมันเป็นตัวแทนของรูปแบบการนำส่งระหว่างสิ่งมีชีวิตและสิ่งไม่มีชีวิต ไวรัสนั้นง่ายมาก อนุภาคของไวรัสแต่ละอนุภาคประกอบด้วย RNA หรือ DNA ที่อยู่ในเปลือกโปรตีนที่เรียกว่า แคปซิด,เรียกว่าอนุภาคติดเชื้อที่ก่อตัวเต็มที่ วิริออน. ไวรัสบางชนิด (เริมหรือไข้หวัดใหญ่) ยังมีซองเพิ่มเติมที่เกิดจากพลาสมาเมมเบรนของเซลล์เจ้าบ้าน ไวรัสสามารถมีชีวิตอยู่และแพร่พันธุ์ได้เฉพาะในเซลล์ของสิ่งมีชีวิตอื่นเท่านั้น ในสภาพแวดล้อมภายนอกพวกเขาไม่แสดงสัญญาณของสิ่งมีชีวิตใด ๆ หลายคนมีรูปแบบของผลึก ขนาดของไวรัสอยู่ระหว่าง 20 ถึง 300 นาโนเมตร
ไวรัสมีโครงสร้างภายในค่อนข้างซับซ้อน แกนกลางของมันมีโมเลกุลกรดนิวคลีอิก (DNA หรือ RNA) หนึ่งโมเลกุล (บางครั้งมากกว่า) กรดนิวคลีอิกของไวรัสที่มีขนาดเล็กที่สุดประกอบด้วยยีน 3-4 ยีน และไวรัสที่ใหญ่ที่สุดมีมากถึง 100 ยีน ภายนอกไวรัสถูกปกคลุมไปด้วย "ฝาครอบ" โปรตีนที่ช่วยปกป้องกรดนิวคลีอิกจากอิทธิพลของสิ่งแวดล้อมที่เป็นอันตราย รูปแบบของไวรัสมีความหลากหลายมาก ขึ้นอยู่กับขนาด ไวรัสแบ่งออกเป็นขนาดใหญ่ (เส้นผ่านศูนย์กลาง 300-400 นาโนเมตร) ขนาดกลาง (80-125 นาโนเมตร) และขนาดเล็ก (20-30 นาโนเมตร) ไวรัสขนาดใหญ่สามารถมองเห็นได้ด้วยกล้องจุลทรรศน์ธรรมดา ในขณะที่ไวรัสที่มีขนาดเล็กกว่านั้นจะถูกศึกษาด้วยกล้องจุลทรรศน์อิเล็กตรอน
มหาวิทยาลัย: มหาวิทยาลัยแห่งรัฐเซาท์อูราล
ปีและเมือง: เชเลียบินสค์ 2554
บทนำ 4
คุณสมบัติของไวรัส ลักษณะของไวรัส5
II โครงสร้างและการจำแนกประเภทของไวรัส 7
III ปฏิกิริยาของไวรัสกับเซลล์ 10
IV ความหมายของไวรัส 12
V โรคไวรัสในมนุษย์ที่มีชื่อเสียงที่สุดบางชนิด 15
VI ไวรัสมีชีวิตอยู่หรือไม่? 17
VII มีชีวิตอยู่ตลอดไป 19
บทสรุป.22
รายการบรรณานุกรม.23
คำอธิบายประกอบ
Fedorova A.V. “ไวรัสเป็นรูปแบบการเปลี่ยนผ่านจากการไม่มีชีวิตไปสู่การมีชีวิต” - Chelyabinsk: SUSU, EiP - 114, 2011, 23 p., 11 ill., 1 table, bibliogr. รายการ - 9 รายการ
บทคัดย่อนี้กล่าวถึงประเด็นต่างๆ เช่น คุณสมบัติและธรรมชาติของไวรัส คุณสมบัติบางประการของไวรัส ได้แก่ มีกรดนิวคลีอิกเพียงชนิดเดียว ไม่มีเมตาบอลิซึมของตัวเอง เป็นต้น โครงสร้างและการจำแนกประเภท ปฏิสัมพันธ์ของไวรัสกับเซลล์ (ไวรัสสามารถมีชีวิตอยู่และแพร่พันธุ์ได้เฉพาะในเซลล์ของสิ่งมีชีวิตอื่นเท่านั้น ภายนอกเซลล์ของสิ่งมีชีวิตพวกมันจะไม่แสดงสัญญาณของสิ่งมีชีวิต) ความสำคัญของไวรัส โรคไวรัสบางชนิด (หัด คางทูม ไข้หวัดใหญ่ โปลิโอ โรคพิษสุนัขบ้า ไข้ทรพิษ ไข้เหลือง ริดสีดวงทวาร โรคไข้สมองอักเสบ มะเร็งบางชนิด (เนื้องอก) โรคเอดส์) มีคำถามว่า “ไวรัสยังมีชีวิตอยู่หรือไม่”
การแนะนำ
ในช่วงปลายศตวรรษที่ผ่านมา ไม่มีใครสงสัยเลยว่าโรคติดเชื้อทุกชนิดมีสาเหตุมาจากจุลินทรีย์ในตัวเอง ซึ่งสามารถต่อสู้กับโรคได้สำเร็จ
“ให้เวลาหน่อยเถอะ” นักวิทยาศาสตร์ด้านแบคทีเรียวิทยากล่าว “แล้วอีกไม่นานก็จะไม่มีโรคเหลืออยู่เลย” แต่หลายปีผ่านไปและคำสัญญาก็ไม่เป็นจริง ประชาชนติดเชื้อโรคหัด โรคปากเท้าเปื่อย โปลิโอ ริดสีดวงทวาร ไข้ทรพิษ ไข้เหลือง และไข้หวัดใหญ่ ผู้คนหลายล้านคนเสียชีวิตจากโรคร้ายแรง แต่ไม่พบจุลินทรีย์ที่ทำให้เกิดโรคดังกล่าว
ในที่สุดในปี พ.ศ. 2435 นักวิทยาศาสตร์ชาวรัสเซีย D.I. Ivanovsky มาถูกทางแล้ว ในขณะที่ศึกษาโรคโมเสกยาสูบ ซึ่งเป็นโรคของใบยาสูบ เขาได้ข้อสรุปว่ามันไม่ได้เกิดจากจุลินทรีย์ แต่เกิดจากสิ่งเล็กๆ น้อยๆ “บางสิ่ง” นี้แทรกซึมผ่านตัวกรองที่บางที่สุดที่สามารถกักเก็บแบคทีเรียได้ ไม่แพร่ขยายในสื่อเทียม จะตายเมื่อถูกความร้อน และไม่สามารถมองเห็นได้ด้วยกล้องจุลทรรศน์แบบใช้แสง พิษกรองได้!
นี่คือข้อสรุปของนักวิทยาศาสตร์ แต่พิษก็คือสาร และสาเหตุของโรคยาสูบก็คือสิ่งมีชีวิต แพร่พันธุ์ได้ดีในใบพืช มาร์ติน วิลเลม ไบรินิค นักพฤกษศาสตร์ชาวเดนมาร์กเรียกไวรัสชนิดใหม่นี้ว่า “บางสิ่ง” โดยเสริมว่าไวรัสนั้นเป็น “ของเหลว มีชีวิต และติดเชื้อได้” แปลจากภาษาละติน "ไวรัส" แปลว่า "พิษ"
ไม่กี่ปีต่อมา F. Leffler และ P. Frosch ค้นพบว่าสาเหตุของโรคปากและเท้าเปื่อย ซึ่งเป็นโรคที่มักพบในปศุสัตว์ก็ผ่านการกรองแบคทีเรียเช่นกัน ในที่สุดในปี 1917 นักแบคทีเรียวิทยาชาวแคนาดา F. de Herelle ค้นพบแบคทีเรียแบคทีเรียซึ่งเป็นไวรัสที่ติดเชื้อแบคทีเรีย
นี่คือวิธีการค้นพบไวรัสของพืช สัตว์ และจุลินทรีย์ เหตุการณ์เหล่านี้เป็นจุดเริ่มต้นของวิทยาศาสตร์ใหม่ - ไวรัสวิทยาศึกษารูปแบบชีวิตที่ไม่ใช่เซลล์
คุณสมบัติของไวรัส ธรรมชาติของไวรัส
2) ไม่มีกระบวนการเผาผลาญของตัวเองและมีเอนไซม์จำนวนจำกัดมาก สำหรับการสืบพันธุ์ จะใช้เมแทบอลิซึมของเซลล์เจ้าบ้าน เอนไซม์ และพลังงานของมัน
ไวรัสไม่ได้แพร่พันธุ์บนอาหารเทียม - พวกมันพิถีพิถันในเรื่องอาหารมากเกินไป น้ำซุปเนื้อธรรมดาที่เหมาะกับแบคทีเรียส่วนใหญ่ไม่เหมาะกับไวรัส . พวกเขาต้องการเซลล์ที่มีชีวิต ไม่ใช่แค่เซลล์ใดๆ แต่ต้องมีการกำหนดไว้อย่างเคร่งครัด เช่นเดียวกับสิ่งมีชีวิตอื่นๆ ไวรัสสามารถแพร่พันธุ์ได้ ไวรัสเป็นกรรมพันธุ์ . ลักษณะทางพันธุกรรมของไวรัสสามารถพิจารณาได้จากช่วงของโฮสต์ที่ได้รับผลกระทบและอาการของโรคที่เกิดขึ้น ตลอดจนความจำเพาะของการตอบสนองทางภูมิคุ้มกันของโฮสต์ตามธรรมชาติหรือสัตว์ทดลองที่ได้รับภูมิคุ้มกันเทียม ผลรวมของลักษณะเหล่านี้ทำให้สามารถระบุคุณสมบัติทางพันธุกรรมของไวรัสได้อย่างชัดเจนและยิ่งไปกว่านั้น - สายพันธุ์ที่มีเครื่องหมายทางพันธุกรรมที่ชัดเจนเช่น neurotropism ของไวรัสไข้หวัดใหญ่บางชนิดเป็นต้น . ความแปรปรวนเป็นอีกด้านหนึ่งของการถ่ายทอดทางพันธุกรรม และในแง่นี้ไวรัสก็เหมือนกับสิ่งมีชีวิตอื่นๆ ที่อาศัยอยู่ในโลกของเรา ในเวลาเดียวกันในไวรัสสามารถสังเกตทั้งความแปรปรวนทางพันธุกรรมที่เกี่ยวข้องกับการเปลี่ยนแปลงของสารพันธุกรรมและความแปรปรวนทางฟีโนไทป์ที่เกี่ยวข้องกับการแสดงออกของจีโนไทป์เดียวกันในสภาวะที่แตกต่างกัน
II โครงสร้างและการจำแนกประเภทของไวรัส
ไม่สามารถมองเห็นไวรัสได้ด้วยกล้องจุลทรรศน์แบบใช้แสง เนื่องจากขนาดของไวรัสมีขนาดเล็กกว่าความยาวคลื่นของแสง สามารถมองเห็นได้โดยใช้กล้องจุลทรรศน์อิเล็กตรอนเท่านั้น
ไวรัสประกอบด้วยส่วนประกอบหลักดังต่อไปนี้
1. แกนกลาง - สารพันธุกรรม (DNA หรือ RNA) ซึ่งมีข้อมูลเกี่ยวกับโปรตีนหลายประเภทที่จำเป็นสำหรับการก่อตัวของไวรัสตัวใหม่
2. เปลือกโปรตีนซึ่งเรียกว่า capsid (จากคำภาษาละติน capsa - box) มักสร้างขึ้นจากหน่วยย่อยที่ซ้ำกันที่เหมือนกัน - แคปโซเมอร์ แคปโซเมอร์สร้างโครงสร้างที่มีความสมมาตรในระดับสูง
3. เมมเบรนไลโปโปรตีนเพิ่มเติม มันถูกสร้างขึ้นจากพลาสมาเมมเบรนของเซลล์เจ้าบ้าน และพบได้ในไวรัสที่มีขนาดค่อนข้างใหญ่เท่านั้น (ไข้หวัดใหญ่ เริม)
แคปซิดและเปลือกเพิ่มเติมมีหน้าที่ป้องกันราวกับปกป้องกรดนิวคลีอิก นอกจากนี้ยังอำนวยความสะดวกในการแทรกซึมของไวรัสเข้าสู่เซลล์ ไวรัสที่มีรูปร่างสมบูรณ์เรียกว่า virion
โครงสร้างแผนผังของไวรัสที่ประกอบด้วย RNA ที่มีสมมาตรแบบขดลวดและซองไลโปโปรตีนเพิ่มเติมจะแสดงทางด้านซ้ายในรูปที่ 1 ส่วนตัดขวางที่ขยายจะแสดงทางด้านขวา
รูปที่ 1 โครงสร้างแผนผังของไวรัส: 1 - คอร์ (RNA แบบเกลียวเดี่ยว); 2 - เปลือกโปรตีน (Capsid); 3 - เมมเบรนไลโปโปรตีนเพิ่มเติม 4 - Capsomeres (ส่วนโครงสร้างของ Capsid)
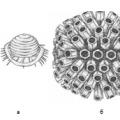
จำนวนแคปโซเมียร์และวิธีการพับจะคงที่อย่างเคร่งครัดสำหรับไวรัสแต่ละประเภท ตัวอย่างเช่น ไวรัสโปลิโอมีแคปโซเมอร์ 32 ตัว และอะดีโนไวรัสมี 252 ตัว
เนื่องจากพื้นฐานของสิ่งมีชีวิตทั้งหมดคือโครงสร้างทางพันธุกรรม ไวรัสจึงถูกจำแนกตามลักษณะของสารทางพันธุกรรม - กรดนิวคลีอิก ไวรัสทั้งหมดแบ่งออกเป็นสองกลุ่มใหญ่ : ไวรัสดีเอ็นเอ(ดีออกซีไวรัส) และ ไวรัสอาร์เอ็นเอ(ไรโบไวรัส). จากนั้นแต่ละกลุ่มจะถูกแบ่งออกเป็นไวรัสกรดนิวคลีอิกแบบเกลียวคู่และเกลียวเดี่ยว เกณฑ์ถัดไปคือประเภทของความสมมาตรของ virions (ขึ้นอยู่กับวิธีการวางแคปโซเมียร์) การมีอยู่หรือไม่มีเปลือกนอกตามเซลล์ - โฮสต์ นอกเหนือจากการจำแนกประเภทเหล่านี้แล้ว ยังมีสิ่งอื่นๆ อีกมากมาย ตัวอย่างเช่นตามประเภทของการแพร่เชื้อจากสิ่งมีชีวิตหนึ่งไปยังอีกสิ่งมีชีวิตหนึ่ง

รูปที่ 2 แผนผังแสดงการจัดเรียงแคปโซเมียร์ในแคปซิดของไวรัส ไวรัสไข้หวัดใหญ่มีลักษณะสมมาตรแบบก้นหอย ลูกบาศก์สมมาตรในไวรัส: เริม - b, อะดีโนไวรัส - ค, โปลิโอเมลิติส - ง
เปลือก.
ไวรัสที่ห่อหุ้มนั้นต่างจากเซลล์ปกติตรงที่ไม่มีความสามารถในการซ่อมแซมเยื่อหุ้มไขมันที่เสียหาย ระบบเสริมสามารถใช้ประโยชน์จากจุดอ่อนนี้ได้เนื่องจากความเสียหายเพียงเล็กน้อยต่อเยื่อหุ้มชั้นนอกจะนำไปสู่การแตกออกและส่งผลให้สูญเสียคุณสมบัติในการติดเชื้อของไวรัส เป็นที่ชัดเจนว่าระบบเสริมซึ่งเป็นส่วนหนึ่งของระบบภูมิคุ้มกันโดยกำเนิด มีบทบาทสำคัญในการต่อสู้กับการติดเชื้อไวรัส อย่างไรก็ตาม MPC จะมีผลกับเชื้อโรคที่มีเยื่อหุ้มไขมันชั้นนอกเท่านั้น แบคทีเรียหรือยีสต์แกรมบวกซึ่งมีผนังเซลล์เพิ่มเติมนั้นไม่สามารถเสริมได้
รูปที่ 3-DNA-โครงสร้างของไวรัส
ก) ไวรัสไข้ทรพิษ
b) เริม - ไวรัส
RNA สายเดี่ยว
c) ไวรัสโรคหัดและคางทูม
d) ไวรัสโรคพิษสุนัขบ้า
e) มะเร็งเม็ดเลือดขาวและไวรัสเอดส์
ไม่มีเปลือก
DNA ที่มีเกลียวคู่
จ) ไอริโด - ไวรัส
g) อะดีโน - ไวรัส
III ปฏิกิริยาระหว่างไวรัสกับเซลล์
ไวรัสสามารถมีชีวิตอยู่และแพร่พันธุ์ได้เฉพาะในเซลล์ของสิ่งมีชีวิตอื่นเท่านั้น ภายนอกเซลล์ของสิ่งมีชีวิต พวกมันจะไม่แสดงสัญญาณของสิ่งมีชีวิตใดๆ ในเรื่องนี้ไวรัสอาจเป็นรูปแบบการพักผ่อนนอกเซลล์ (varion) หรือรูปแบบการทำซ้ำภายในเซลล์ - พืช Varis แสดงให้เห็นถึงความมีชีวิตที่ยอดเยี่ยม โดยเฉพาะอย่างยิ่งสามารถทนต่อแรงกดดันได้สูงถึง 6,000 atm และทนต่อรังสีในปริมาณสูง แต่จะตายที่อุณหภูมิสูง การฉายรังสีด้วยรังสียูวี ตลอดจนการสัมผัสกับกรดและยาฆ่าเชื้อ
ปฏิสัมพันธ์ของไวรัสกับเซลล์ต้องผ่านหลายขั้นตอนตามลำดับ
1.ขั้นแรกคือ การดูดซับความแปรผันบนพื้นผิวของเซลล์เป้าหมายซึ่งเพื่อการนี้จะต้องมีตัวรับพื้นผิวที่เหมาะสม มันอยู่กับพวกเขาว่าอนุภาคของไวรัสมีปฏิกิริยากันโดยเฉพาะหลังจากนั้นพวกมันก็ถูกผูกไว้อย่างแน่นหนาด้วยเหตุนี้เซลล์จึงไม่ไวต่อไวรัสทุกชนิด นี่คือสิ่งที่อธิบายความแน่นอนที่เข้มงวดของเส้นทางการแทรกซึมของไวรัส ตัวอย่างเช่น เซลล์ในเยื่อเมือกของระบบทางเดินหายใจมีตัวรับไวรัสไข้หวัดใหญ่ แต่เซลล์ผิวหนังไม่มี ดังนั้นจึงคุณไม่สามารถแพร่เชื้อไข้หวัดใหญ่ทางผิวหนังได้ - ต้องสูดอนุภาคของไวรัสในอากาศ ไวรัสตับอักเสบ A หรือ B จะแทรกซึมและเพิ่มจำนวนเฉพาะในเซลล์ตับ และไวรัสคางทูม (คางทูม) - ในเซลล์ของต่อมน้ำลายหู ฯลฯ
2. ระยะที่ 2 ประกอบด้วย การเจาะความแปรปรวนทั้งหมดหรือกรดนิวคลีอิกเข้าไปในเซลล์เจ้าบ้าน
3. ขั้นที่สามเรียกว่า ลดโปรตีน. ในระหว่างกระบวนการนี้ พาหะของข้อมูลทางพันธุกรรมของไวรัสซึ่งก็คือกรดนิวคลีอิกของไวรัสจะถูกปล่อยออกมา
4. ในช่วงระยะที่ 4 ขึ้นอยู่กับกรดนิวคลีอิกของไวรัส การสังเคราะห์สารประกอบที่จำเป็นสำหรับไวรัส
5. ในขั้นที่ 5 เกิดขึ้น การสังเคราะห์ส่วนประกอบของอนุภาคไวรัส- กรดนิวคลีอิกและโปรตีนแคปซิด และส่วนประกอบทั้งหมดถูกสังเคราะห์หลายครั้ง
6. ในช่วงระยะที่ 6 จากการสังเคราะห์กรดนิวคลีอิกและโปรตีนจำนวนมากก่อนหน้านี้ virions ใหม่เกิดขึ้นจากการประกอบตัวเอง
7. ขั้นตอนสุดท้าย - ขั้นตอนที่เจ็ด - หมายถึงการปล่อยอนุภาคไวรัสที่รวมตัวกันใหม่ออกจากเซลล์เจ้าบ้าน กระบวนการนี้เกิดขึ้นแตกต่างกันไปสำหรับไวรัสแต่ละชนิด ในไวรัสบางชนิดสิ่งนี้จะมาพร้อมกับการตายของเซลล์เนื่องจากการปลดปล่อยเอนไซม์ lytic ในไลโซโซม - การสลายเซลล์ในกรณีอื่นๆ การแปรผันจะออกจากเซลล์ที่มีชีวิตโดยการแตกหน่อ แต่แม้ในกรณีนี้ เซลล์จะตายไปตามกาลเวลา
IV ความสำคัญของไวรัส
วิทยาศาสตร์รู้จักไวรัสของแบคทีเรีย พืช แมลง สัตว์ และมนุษย์ มีมากกว่า 1,000 ตัว การแพร่พันธุ์ของไวรัสที่เกี่ยวข้องกับการทำลายเซลล์ทำให้เกิดอาการเจ็บปวดในร่างกาย ไวรัสทำให้เกิดโรคในมนุษย์หลายชนิด เช่น โรคหัด คางทูม ไข้หวัดใหญ่ โปลิโอ โรคพิษสุนัขบ้า ไข้ทรพิษ ไข้เหลือง ริดสีดวงทวาร โรคไข้สมองอักเสบ มะเร็งบางชนิด (เนื้องอก) โรคเอดส์ ไม่ใช่เรื่องแปลกที่คนเราจะเริ่มมีหูด ทุกคนรู้ดีว่าหลังจากเป็นหวัดพวกเขามักจะ "กวาด" ริมฝีปากและปีกจมูกอย่างไร สิ่งเหล่านี้ล้วนเป็นโรคไวรัสเช่นกัน นักวิทยาศาสตร์พบว่าไวรัสจำนวนมากอาศัยอยู่ในร่างกายมนุษย์ แต่ก็ไม่ได้แสดงออกมาเสมอไป มีเพียงร่างกายที่อ่อนแอเท่านั้นที่ไวต่อผลกระทบของไวรัสที่ทำให้เกิดโรค เส้นทางการติดเชื้อไวรัสนั้นแตกต่างกันมาก: ผ่านผิวหนังผ่านแมลงและเห็บกัด; ผ่านทางน้ำลาย น้ำมูก และสารคัดหลั่งอื่น ๆ ของผู้ป่วย ผ่านอากาศ กับอาหาร; ทางเพศและอื่น ๆ
การติดเชื้อแบบหยดเป็นวิธีการแพร่กระจายโรคระบบทางเดินหายใจที่พบบ่อยที่สุด การไอและจามจะปล่อยหยดของเหลว (เมือกและน้ำลาย) จำนวนนับล้านหยดไปในอากาศ ละอองเหล่านี้รวมถึงจุลินทรีย์ที่มีชีวิตที่มีอยู่สามารถสูดดมโดยผู้อื่นได้ ในสัตว์ ไวรัสทำให้เกิดโรคปากและเท้าเปื่อย โรคระบาด และโรคพิษสุนัขบ้า ในแมลง - polyhedrosis, granulomatosis; ในพืช - โมเสกหรือการเปลี่ยนแปลงสีของใบไม้หรือดอกไม้อื่น ๆ การม้วนงอของใบและการเปลี่ยนแปลงรูปร่างอื่น ๆ การแคระแกร็น ในที่สุดในแบคทีเรีย - การสลายตัวของพวกเขา แนวคิดของไวรัสในฐานะ "ผู้ทำลาย" ที่ไม่หยุดนิ่งในการศึกษาไวรัสกลุ่มพิเศษที่ติดเชื้อแบคทีเรีย เรากำลังพูดถึงแบคทีเรีย ความสามารถของฟาจในการทำลายแบคทีเรียสามารถใช้เพื่อรักษาโรคบางชนิดที่เกิดจากแบคทีเรียเหล่านี้ได้ ฟาจส์กลายเป็นไวรัสกลุ่มแรกที่ "เชื่อง" โดยมนุษย์อย่างแท้จริง พวกเขาจัดการกับเพื่อนบ้านที่ใกล้ที่สุดในพิภพเล็ก ๆ อย่างรวดเร็วและไร้ความปราณี โรคระบาด ไทฟอยด์ โรคบิด และอหิวาตกโรค วิบริโอ “ละลาย” ต่อหน้าต่อตาเราหลังจากพบกับไวรัสเหล่านี้ เริ่มใช้เพื่อป้องกันและรักษาโรคติดเชื้อหลายชนิด แต่น่าเสียดายที่ความสำเร็จครั้งแรกตามมาด้วยความล้มเหลว เนื่องจากในร่างกายมนุษย์ ฟาจไม่ได้โจมตีแบคทีเรียอย่างแข็งขันเหมือนกับในหลอดทดลอง นอกจากนี้แบคทีเรียยังกลายเป็น "เจ้าเล่ห์" มากกว่าศัตรู: พวกมันปรับตัวเข้ากับฟาจอย่างรวดเร็วและไม่ไวต่อการกระทำของพวกมัน
หลังจากการค้นพบยาปฏิชีวนะ ฟาจก็ถอยกลับไปเป็นยา แต่ยังคงใช้ในการจดจำแบคทีเรียได้สำเร็จ ความจริงก็คือฟาจสามารถค้นหา “แบคทีเรียของพวกมัน” ได้อย่างแม่นยำมากและละลายพวกมันได้อย่างรวดเร็ว คุณสมบัติที่คล้ายกันของฟาจเป็นพื้นฐานสำหรับการวินิจฉัยการรักษา โดยปกติจะทำดังนี้: แบคทีเรียที่แยกได้จากร่างกายของผู้ป่วยจะเติบโตบนอาหารที่เป็นของแข็ง หลังจากนั้นฟาจต่างๆ เช่น โรคบิดและไทฟอยด์ จะถูกนำไปใช้กับ "สนามหญ้า" ที่เกิดขึ้น หลังจากผ่านไปหนึ่งวัน จานจะถูกตรวจสอบภายใต้แสง และพิจารณาว่าฟาจใดที่ทำให้เกิดการละลายของแบคทีเรีย หากฟาจโรคบิดมีผลเช่นนั้น แบคทีเรียโรคบิดจะถูกแยกออกจากร่างกายของผู้ป่วย ถ้าไทฟอยด์ แบคทีเรียไทฟอยด์จะถูกแยกออก
บางครั้งไวรัสที่แพร่ระบาดในสัตว์และแมลงก็เข้ามาช่วยเหลือมนุษย์ได้ กว่ายี่สิบปีที่แล้วในออสเตรเลีย ปัญหาการต่อสู้กับกระต่ายป่าเริ่มรุนแรงขึ้น จำนวนสัตว์ฟันแทะเหล่านี้มีจำนวนถึงสัดส่วนที่น่าตกใจ พวกเขาทำลายพืชผลเร็วกว่าตั๊กแตนและกลายเป็นหายนะระดับชาติอย่างแท้จริง วิธีการทั่วไปในการจัดการกับพวกมันกลับกลายเป็นว่าไม่ได้ผล จากนั้นนักวิทยาศาสตร์ก็ปล่อยไวรัสชนิดพิเศษเพื่อต่อสู้กับกระต่าย ซึ่งสามารถทำลายสัตว์ที่ติดเชื้อได้เกือบทั้งหมด แต่จะแพร่กระจายโรคนี้ไปยังกระต่ายที่ขี้อายและระมัดระวังได้อย่างไร? ยุงก็ช่วย พวกเขาเล่นบทบาทเป็น "เข็มบิน" แพร่ไวรัสจากกระต่ายสู่กระต่าย ในขณะเดียวกัน ยุงก็ยังมีสุขภาพแข็งแรงสมบูรณ์
มีตัวอย่างอื่นๆ ของความสำเร็จในการใช้ไวรัสเพื่อฆ่าแมลงศัตรูพืช ทุกคนรู้ถึงความเสียหายที่เกิดจากหนอนผีเสื้อและแมลงวัน ครั้งแรกกินใบของพืชที่มีประโยชน์ส่วนหลังติดเชื้อต้นไม้ในสวนและป่าไม้ พวกเขากำลังต่อสู้กับไวรัสที่เรียกว่า polyhedrosis และ granulosis ซึ่งถูกพ่นในพื้นที่ขนาดเล็กด้วยอะตอมไมเซอร์ และใช้เครื่องบินเพื่อรักษาพื้นที่ขนาดใหญ่ สิ่งนี้เกิดขึ้นในสหรัฐอเมริกาเมื่อต่อสู้กับหนอนผีเสื้อที่แพร่ระบาดในทุ่งหญ้าชนิตและในแคนาดาเมื่อทำลายแมลงวันสน
จะเกิดอะไรขึ้นกับเซลล์หากไม่ได้ติดไวรัสเพียงตัวเดียว แต่มีไวรัสถึงสองตัว? หากคุณตัดสินใจว่าในกรณีนี้ โรคของเซลล์จะแย่ลงและการตายของเซลล์จะเร็วขึ้น แสดงว่าคุณคิดผิด ปรากฎว่าการมีไวรัสตัวหนึ่งอยู่ในเซลล์มักจะปกป้องไวรัสจากผลการทำลายล้างของไวรัสตัวอื่นได้อย่างน่าเชื่อถือ ปรากฏการณ์นี้เรียกว่าการรบกวนของไวรัสโดยนักวิทยาศาสตร์ มีความเกี่ยวข้องกับการผลิตโปรตีนพิเศษ - อินเตอร์เฟอรอนซึ่งในเซลล์จะกระตุ้นกลไกการป้องกันที่สามารถแยกแยะไวรัสออกจากไวรัสและเลือกยับยั้งไวรัสได้ อินเตอร์เฟอรอนยับยั้งการแพร่พันธุ์ของไวรัสส่วนใหญ่ในเซลล์ ปัจจุบันอินเตอร์เฟอรอนที่ผลิตขึ้นเพื่อเป็นยารักษาโรคถูกนำมาใช้ในการรักษาและป้องกันโรคไวรัสหลายชนิด
V โรคไวรัสบางชนิดที่มนุษย์รู้จักมากที่สุด
ไข้หวัดใหญ่ยังคงเป็น “ราชา” ของโรคระบาด ไม่มีโรคใดสามารถเข้าถึงผู้คนหลายร้อยล้านคนได้ในเวลาอันสั้น และผู้คนมากกว่าหนึ่งพันล้านคนป่วยด้วยไข้หวัดใหญ่ในช่วงที่มีการระบาดใหญ่! ไม่เพียงแต่เกิดขึ้นในช่วงที่มีการระบาดใหญ่ที่น่าจดจำในปี 1918 เท่านั้น แต่ยังเกิดขึ้นเมื่อไม่นานมานี้ ในปี 1957 ซึ่งเป็นช่วงที่ไข้หวัดใหญ่ "เอเชีย" ระบาด และในปี 1968 เมื่อไข้หวัดใหญ่ "ฮ่องกง" ปรากฏขึ้น รู้จักไวรัสไข้หวัดใหญ่หลายชนิด - A, B, C เป็นต้น ภายใต้อิทธิพลของปัจจัยด้านสิ่งแวดล้อมจำนวนอาจเพิ่มขึ้น เนื่องจากภูมิคุ้มกันต่อไข้หวัดใหญ่อยู่ในระยะสั้นและเฉพาะเจาะจง การเจ็บป่วยซ้ำๆ จึงเป็นไปได้ในฤดูกาลเดียว จากสถิติพบว่าประชากรป่วยไข้หวัดใหญ่โดยเฉลี่ยประมาณ 20-35% ทุกปี
ไข้ทรพิษเป็นหนึ่งในโรคที่เก่าแก่ที่สุด คำอธิบายของไข้ทรพิษพบในกระดาษปาปิรัสอียิปต์ของ Amenophis I ซึ่งรวบรวมเมื่อ 4,000 ปีก่อนคริสตกาล รอยโรคไข้ทรพิษถูกเก็บรักษาไว้บนผิวหนังของมัมมี่ที่ถูกฝังในอียิปต์เมื่อ 3,000 ปีก่อนคริสตกาล การกล่าวถึงไข้ทรพิษซึ่งชาวจีนเรียกว่า "พิษจากอกแม่" มีอยู่ในแหล่งที่มาของจีนที่เก่าแก่ที่สุด - บทความ "Cheu-Cheufa" (1120 ปีก่อนคริสตกาล) คำอธิบายคลาสสิกครั้งแรกของไข้ทรพิษได้รับจากแพทย์ชาวอาหรับ Rhazes
ตารางที่ 1-ลักษณะของโรค
โรคเอดส์เป็นโรคติดเชื้อชนิดใหม่ซึ่งผู้เชี่ยวชาญยอมรับว่าเป็นโรคระบาดระดับโลกอย่างแท้จริงครั้งแรกในประวัติศาสตร์ของมนุษย์ ทั้งโรคระบาด ไข้ทรพิษ หรืออหิวาตกโรคก็ไม่เคยเกิดขึ้นมาก่อน เนื่องด้วยโรคเอดส์ไม่เหมือนกับโรคเหล่านี้และโรคอื่นๆ ในมนุษย์อย่างแน่นอน
|
ชื่อโรค |
เชื้อโรค |
พื้นที่ของร่างกายได้รับผลกระทบ |
วิธีการจัดจำหน่าย |
ประเภทของการฉีดวัคซีน |
|
Myxovirus หนึ่งในสามประเภท - A, B และ C - โดยมีระดับความรุนแรงที่แตกต่างกัน |
ระบบทางเดินหายใจ: เยื่อบุผิวเยื่อบุหลอดลมและหลอดลม |
การติดเชื้อแบบหยด |
ไวรัสที่ถูกฆ่า: สายพันธุ์ของไวรัสที่ถูกฆ่าจะต้องตรงกับสายพันธุ์ของไวรัสที่ทำให้เกิด ความต่อเนื่องของตารางที่ 1 โรค |
|
|
เย็น |
ไวรัสหลากหลายชนิด ส่วนใหญ่มักเป็นไรโนไวรัส (ไวรัสที่มี RNA) |
ทางเดินหายใจ: โดยปกติจะเป็นส่วนบนเท่านั้น |
การติดเชื้อแบบหยด |
ไวรัสที่มีชีวิตหรือไม่มีการใช้งานจะได้รับการบริหารโดยการฉีดเข้ากล้าม การฉีดวัคซีนไม่ได้ผลมากนักเนื่องจากมีไรโนไวรัสหลายสายพันธุ์ |
|
ไวรัส Variola (ไวรัสที่มี DNA) หนึ่งในไวรัสไข้ทรพิษ |
แอร์เวย์แล้วผิวหนัง |
การติดเชื้อแบบหยด (สามารถติดต่อผ่านบาดแผลทางผิวหนังได้) |
ไวรัสที่มีชีวิตอ่อนแอ (ลดทอน) จะถูกนำเข้าสู่รอยขีดข่วนบนผิวหนัง ยังไม่ได้ใช้ในปัจจุบัน |
|
|
คางทูม (paratitis) |
ระบบทางเดินหายใจ จากนั้นจึงเกิดการติดเชื้อทั่วร่างกายผ่านทางเลือด ต่อมน้ำลายได้รับผลกระทบเป็นพิเศษ และในผู้ชายที่เป็นผู้ใหญ่ก็ส่งผลต่ออัณฑะด้วย |
การติดเชื้อแบบหยด (หรือการแพร่เชื้อทางปากด้วยน้ำลายติดเชื้อ) |
ไวรัสลดทอนสด |
|
|
Paramyxovirus (RNA - มีไวรัส) |
ทางเดินหายใจ (จากช่องปากถึงหลอดลม) จากนั้นผ่านไปยังผิวหนังและลำไส้ |
การติดเชื้อแบบหยด |
ไวรัสลดทอนสด |
|
|
โรคหัดเยอรมัน (หัดเยอรมัน) |
ไวรัสหัดเยอรมัน |
สายการบิน ต่อมน้ำเหลืองที่ปากมดลูก ดวงตา และผิวหนัง |
การติดเชื้อแบบหยด |
ไวรัสลดทอนสด |
|
โปลิโอไมเอลิติส (อัมพาตในทารก) |
โปลิโอไวรัส (picornavirus; ไวรัสที่มี RNA รู้จักสามสายพันธุ์) |
คอและลำไส้จากนั้นเป็นเลือด บางครั้งเซลล์ประสาทสั่งการของไขสันหลังก็อาจเกิดอัมพาตได้ |
การติดเชื้อแบบหยดหรือผ่านอุจจาระของมนุษย์ |
ไวรัสเชื้อเป็นจะถูกให้ทางปาก โดยปกติจะรับประทานบนน้ำตาลก้อน |
VI ไวรัสมีชีวิตอยู่หรือไม่?
พิจารณาสองมุมมอง:
- ถ้าเราพิจารณาโครงสร้างสิ่งมีชีวิตที่มีกรดนิวคลีอิกและสามารถสืบพันธุ์ได้เอง เราก็สามารถยอมรับมุมมองที่ว่าไวรัสมีชีวิตได้
- หากเราสันนิษฐานว่ามีเพียงโครงสร้างที่มีโครงสร้างเซลล์เท่านั้นที่ยังมีชีวิตอยู่ ไวรัสก็เป็นสสารที่ไม่มีชีวิต (โพลีเมอร์)
A. Lehninger ใน “ชีวเคมี” ถือว่าไวรัสเป็นโครงสร้างที่ยืนอยู่บนธรณีประตูของชีวิตและเป็นตัวแทนของสารเชิงซ้อนซูปราโมเลคิวลาร์ที่เสถียรซึ่งประกอบด้วยโมเลกุลกรดนิวคลีอิกและหน่วยย่อยโปรตีนจำนวนมาก จัดเรียงตามลำดับที่แน่นอนและสร้างโครงสร้างสามมิติที่เฉพาะเจาะจง ในบรรดาคุณสมบัติที่สำคัญที่สุดของไวรัสเขาตั้งข้อสังเกต:
- ไม่สามารถสืบพันธุ์ได้เองในรูปของยาบริสุทธิ์
- ความสามารถในการควบคุมการจำลองแบบ (เซลล์ที่ติดเชื้อ);
- ไวรัสหลากหลายขนาด รูปร่าง และองค์ประกอบทางเคมี
ไวรัสตั้งอยู่บนขอบเขตระหว่างสิ่งมีชีวิตและไม่มีชีวิต สิ่งนี้บ่งชี้ถึงการมีอยู่ของสเปกตรัมที่ต่อเนื่องของโลกอินทรีย์ที่ซับซ้อนมากขึ้น ซึ่งเริ่มต้นด้วยโมเลกุลธรรมดา ๆ และจบลงด้วยระบบเซลล์ที่ซับซ้อนที่สุด
หินเช่นเดียวกับหยดของเหลวที่เกิดกระบวนการเผาผลาญ แต่ไม่มีสารพันธุกรรมและไม่สามารถสืบพันธุ์ได้เองนั้นเป็นวัตถุไม่มีชีวิตอย่างไม่ต้องสงสัย แบคทีเรียเป็นสิ่งมีชีวิตและถึงแม้ว่ามันจะประกอบด้วยเซลล์เดียว แต่ก็สามารถผลิตพลังงานและสังเคราะห์สารที่รับประกันการดำรงอยู่และการสืบพันธุ์ของมัน จะพูดอะไรเกี่ยวกับเมล็ดพืชในบริบทนี้ได้บ้าง ไม่ใช่ทุกเมล็ดที่แสดงสัญญาณแห่งชีวิต อย่างไรก็ตาม เมื่ออยู่นิ่ง ก็มีศักยภาพที่ได้รับจากสิ่งมีชีวิตอย่างไม่ต้องสงสัย และสามารถรับรู้ได้ภายใต้เงื่อนไขบางประการ ในเวลาเดียวกัน เมล็ดพืชสามารถถูกทำลายอย่างถาวรได้ และจากนั้นศักยภาพก็จะยังไม่เกิดขึ้นจริง ในเรื่องนี้ไวรัสนั้นชวนให้นึกถึงเมล็ดพันธุ์มากกว่าเซลล์ที่มีชีวิต: มีความสามารถบางอย่างที่อาจไม่เป็นจริง แต่ไม่มีความสามารถในการดำรงอยู่โดยอัตโนมัติ
ทั้งยีนหรือโปรตีนของเซลล์หรือไวรัสนั้นทำหน้าที่เป็นสารที่มีชีวิต และเซลล์ที่ไม่มีนิวเคลียสก็คล้ายกับบุคคลที่ถูกตัดหัวเนื่องจากไม่มีความซับซ้อนในระดับวิกฤต ไวรัสก็ไม่สามารถเข้าถึงระดับนี้ได้ ดังนั้นชีวิตจึงสามารถนิยามได้ว่าเป็นสภาวะฉุกเฉินที่ซับซ้อน ซึ่งรวมถึง "องค์ประกอบพื้นฐาน" พื้นฐานเดียวกันกับที่ไวรัสครอบครอง หากเราปฏิบัติตามตรรกะนี้ ไวรัสซึ่งไม่ใช่วัตถุที่มีชีวิตในความหมายที่เข้มงวดของคำนี้ ยังคงไม่สามารถจัดเป็นระบบเฉื่อยได้ พวกมันอยู่บนขอบเขตระหว่างสิ่งมีชีวิตและไม่มีชีวิต
VII มีชีวิตอยู่ตลอดไป
ไวรัสที่อยู่ในตำแหน่งกึ่งกลางระหว่างสิ่งมีชีวิตและไม่มีชีวิตแสดงคุณสมบัติที่ไม่คาดคิด นี่คือหนึ่งในนั้น โดยทั่วไปแล้ว ไวรัสจะแพร่พันธุ์เฉพาะในเซลล์ที่มีชีวิตเท่านั้น แต่พวกมันยังสามารถเติบโตในเซลล์ที่ตายแล้ว และบางครั้งก็ทำให้เซลล์หลังกลับมามีชีวิตอีกครั้งด้วยซ้ำ น่าประหลาดใจที่ไวรัสบางชนิดเมื่อถูกทำลายสามารถเกิดใหม่เป็น "ชีวิตยืม" ได้
เซลล์ที่ DNA นิวเคลียร์ถูกทำลายนั้น "ตาย" อย่างแท้จริง: ปราศจากสารพันธุกรรมพร้อมคำแนะนำในการทำกิจกรรม แต่ไวรัสสามารถใช้ส่วนประกอบของเซลล์และไซโตพลาสซึมที่เหลืออยู่ในการจำลองได้ มันควบคุมอุปกรณ์ของเซลล์และบังคับให้ใช้ยีนของไวรัสเป็นแหล่งคำแนะนำในการสังเคราะห์โปรตีนของไวรัสและการจำลองจีโนมของไวรัส ความสามารถเฉพาะตัวของไวรัสในการพัฒนาในเซลล์ที่ตายแล้วนั้นแสดงให้เห็นได้ชัดเจนที่สุดเมื่อโฮสต์เป็นสิ่งมีชีวิตเซลล์เดียว โดยหลักแล้วพวกมันอาศัยอยู่ในมหาสมุทร
รูปที่ 4 ไวรัสโมเสกยาสูบ
แบคทีเรีย ไซยาโนแบคทีเรียสังเคราะห์แสง และสาหร่าย ซึ่งเป็นโฮสต์ของไวรัสในทะเล มักถูกฆ่าด้วยรังสีอัลตราไวโอเลต ซึ่งทำลาย DNA ของพวกมัน ในเวลาเดียวกัน ไวรัสบางชนิด (“ที่อยู่อาศัย” ของสิ่งมีชีวิต) จะเปิดกลไกการสังเคราะห์เอนไซม์ที่จะฟื้นฟูโมเลกุลที่เสียหายของเซลล์เจ้าบ้านและนำมันกลับมามีชีวิตอีกครั้ง ตัวอย่างเช่น ไซยาโนแบคทีเรียมีเอนไซม์ที่เกี่ยวข้องกับการสังเคราะห์ด้วยแสง และเมื่อสัมผัสกับแสงที่มากเกินไป บางครั้งมันก็ถูกทำลาย ส่งผลให้เซลล์ตาย จากนั้นไวรัสที่เรียกว่าไซยาโนฟาจจะ "เปิด" ซึ่งเป็นการสังเคราะห์อะนาล็อกของเอนไซม์สังเคราะห์แสงจากแบคทีเรียซึ่งมีความทนทานต่อรังสียูวีได้ดีกว่า หากไวรัสดังกล่าวแพร่เชื้อไปยังเซลล์ที่ตายแล้ว เอนไซม์สังเคราะห์แสงสามารถทำให้เซลล์กลับมามีชีวิตได้ ดังนั้นไวรัสจึงมีบทบาทเป็น "เครื่องช่วยชีวิตด้วยยีน"
การได้รับรังสี UV ในปริมาณที่มากเกินไปอาจทำให้ไซยาโนฟาจตายได้ แต่บางครั้งพวกมันสามารถกลับมามีชีวิตอีกครั้งได้ด้วยการซ่อมแซมหลายครั้ง โดยปกติแล้วจะมีไวรัสหลายตัวอยู่ในเซลล์เจ้าบ้านแต่ละเซลล์ และหากได้รับความเสียหาย ก็สามารถประกอบจีโนมของไวรัสทีละชิ้นได้ ส่วนต่างๆ ของจีโนมสามารถทำหน้าที่เป็นซัพพลายเออร์ของยีนแต่ละตัวได้ ซึ่งเมื่อรวมกับยีนอื่นๆ จะช่วยฟื้นฟูการทำงานของจีโนมได้อย่างสมบูรณ์โดยไม่ต้องสร้างไวรัสทั้งหมด ไวรัสเป็นสิ่งมีชีวิตชนิดเดียวที่สามารถเกิดใหม่จากเถ้าถ่านได้เช่นเดียวกับนกฟีนิกซ์
เราร่วมกับเพื่อนร่วมงานที่สถาบันวิทยาศาสตร์สุขภาพแห่งมหาวิทยาลัยออริกอนสำหรับวัคซีนและการบำบัดด้วยยีน เราเสนอว่ามีวิธีที่สาม: ยีนในตอนแรกมีต้นกำเนิดของไวรัส แต่ต่อมาได้ตั้งอาณานิคมของสิ่งมีชีวิตสองสายเลือดที่แตกต่างกัน เช่น แบคทีเรียและสัตว์มีกระดูกสันหลัง ยีนที่แบคทีเรียมอบให้มนุษยชาติสามารถถ่ายทอดไปยังเชื้อสายทั้งสองที่ไวรัสกล่าวถึงได้
รูปที่ 5. ไวรัสลิ้นสีน้ำเงิน
นอกจากนี้เรายังมั่นใจว่านิวเคลียสของเซลล์นั้นมีต้นกำเนิดจากไวรัส การปรากฏตัวของนิวเคลียสไม่สามารถอธิบายได้ด้วยการปรับตัวของสิ่งมีชีวิตโปรคาริโอตอย่างค่อยเป็นค่อยไปให้เข้ากับสภาวะที่เปลี่ยนแปลงไป มันสามารถถูกสร้างขึ้นบนพื้นฐานของ DNA ของไวรัสที่มีน้ำหนักโมเลกุลสูงที่มีอยู่ก่อน ซึ่งได้สร้าง "บ้าน" ถาวรสำหรับตัวมันเองภายในเซลล์โปรคาริโอต สิ่งนี้ได้รับการยืนยันจากข้อเท็จจริงที่ว่ายีน DNA polymerase ของ phage T4 (ฟาจคือไวรัสที่ติดเชื้อแบคทีเรีย) นั้นมีลำดับนิวคลีโอไทด์ใกล้เคียงกับยีน DNA polymerase ของทั้งยูคาริโอตและไวรัสที่ติดเชื้อ นอกจากนี้ Patrick Fortere จากมหาวิทยาลัย Paris South ซึ่งศึกษาเอนไซม์ที่เกี่ยวข้องกับการจำลองดีเอ็นเอ ได้ข้อสรุปว่ายีนที่กำหนดการสังเคราะห์ในยูคาริโอตนั้นมีต้นกำเนิดจากไวรัส
ไวรัสส่งผลกระทบต่อสิ่งมีชีวิตทุกรูปแบบบนโลกอย่างแน่นอน และมักจะกำหนดชะตากรรมของพวกมัน ในขณะเดียวกันก็พัฒนาไปด้วย หลักฐานโดยตรงมาจากการเกิดขึ้นของไวรัสใหม่ๆ เช่น ไวรัสภูมิคุ้มกันบกพร่องในมนุษย์ (HIV) ซึ่งเป็นสาเหตุของโรคเอดส์
ไวรัสปรับเปลี่ยนขอบเขตระหว่างโลกทางชีววิทยาและชีวเคมีอยู่ตลอดเวลา ยิ่งเราก้าวหน้าในการศึกษาจีโนมของสิ่งมีชีวิตต่างๆ มากเท่าไร เราก็จะพบหลักฐานมากขึ้นเกี่ยวกับการมีอยู่ของยีนจากแหล่งน้ำที่เก่าแก่และมีชีวิตชีวามากเท่านั้น ผู้ได้รับรางวัลโนเบล Salvador Luria พูดถึงอิทธิพลของไวรัสที่มีต่อวิวัฒนาการในปี 1969: “บางทีไวรัสที่มีความสามารถในการเข้าและออกจากจีโนมของเซลล์อาจเป็นผู้มีส่วนร่วมในกระบวนการเพิ่มประสิทธิภาพสารพันธุกรรมของสิ่งมีชีวิตทั้งหมดในระหว่างการวิวัฒนาการ เพียงแค่ เราไม่ได้สังเกตเห็นมัน” ไม่ว่าโลกที่มีชีวิตหรือไม่มีชีวิตจะเป็นเช่นไร ถึงเวลาที่ต้องพิจารณาว่าไวรัสไม่ได้อยู่อย่างโดดเดี่ยว แต่ต้องคำนึงถึงความเชื่อมโยงอย่างต่อเนื่องกับสิ่งมีชีวิตด้วย
บทสรุป
การต่อสู้กับการติดเชื้อไวรัสนั้นเกี่ยวข้องกับความยากลำบากมากมายโดยเฉพาะอย่างยิ่งควรสังเกตภูมิคุ้มกันของไวรัสต่อยาปฏิชีวนะเป็นพิเศษ ไวรัสกำลังกลายพันธุ์อย่างแข็งขันและมีสายพันธุ์ใหม่ปรากฏขึ้นเป็นประจำซึ่งยังไม่พบ "อาวุธ" ประการแรก สิ่งนี้ใช้ได้กับไวรัส RNA ซึ่งจีโนมมักจะมีขนาดใหญ่กว่าและมีความเสถียรน้อยกว่า จนถึงปัจจุบันการต่อสู้กับการติดเชื้อไวรัสจำนวนมากกำลังพัฒนาเพื่อประโยชน์ของมนุษย์ส่วนใหญ่เกิดจากการฉีดวัคซีนสากลของประชากรเพื่อวัตถุประสงค์ในการป้องกัน ในที่สุดเหตุการณ์ดังกล่าวนำไปสู่ความจริงที่ว่าตามที่ผู้เชี่ยวชาญระบุว่าไวรัสไข้ทรพิษได้หายไปจากธรรมชาติแล้ว อันเป็นผลมาจากการฉีดวัคซีนสากลในประเทศของเราเมื่อปี พ.ศ. 2504 โรคระบาดโปลิโอถูกกำจัดให้หมดไป อย่างไรก็ตาม ธรรมชาติยังคงทดสอบมนุษย์เป็นครั้งคราว โดยนำเสนอความประหลาดใจในรูปแบบของไวรัสชนิดใหม่ที่ก่อให้เกิดโรคร้ายแรง ตัวอย่างที่เด่นชัดที่สุดคือไวรัสโรคภูมิคุ้มกันบกพร่องของมนุษย์ ซึ่งเป็นการต่อสู้กับมนุษย์ที่ยังคงพ่ายแพ้อยู่ การแพร่กระจายของมันสอดคล้องกับการระบาดใหญ่อยู่แล้ว
สำคัญ! บทคัดย่อที่นำเสนอทั้งหมดให้ดาวน์โหลดฟรีมีจุดมุ่งหมายเพื่อจัดทำแผนหรือพื้นฐานสำหรับงานทางวิทยาศาสตร์ของคุณเอง
เพื่อน! คุณมีโอกาสพิเศษที่จะช่วยเหลือนักเรียนเช่นเดียวกับคุณ! หากเว็บไซต์ของเราช่วยให้คุณหางานที่คุณต้องการได้ คุณจะเข้าใจอย่างแน่นอนว่างานที่คุณเพิ่มเข้าไปจะทำให้งานของผู้อื่นง่ายขึ้นได้อย่างไร
ในความเห็นของคุณ หากบทคัดย่อมีคุณภาพไม่ดี หรือคุณเคยเห็นงานนี้แล้ว โปรดแจ้งให้เราทราบ
เชื่อกันว่าไวรัส (จาก Lat. ไวรัส- ยาพิษ) - สิ่งที่น่ารังเกียจซึ่งไม่ได้นำมาซึ่งอะไรนอกจากปัญหา แต่นี่เป็นความผิดพลาดร้ายแรง ไวรัสเป็นผู้สร้างธรรมชาติของสิ่งมีชีวิตและเป็นกลไกสำคัญในการวิวัฒนาการของมัน
ข้อโต้แย้งหลักประการหนึ่งที่ขัดแย้งกับสมมติฐานที่ว่าไวรัส "หลบหนี" ออกจากเซลล์ก็คือความจริงที่ว่าระบบพันธุกรรมของไวรัสมีความหลากหลายมากกว่าระบบทางพันธุกรรมในเซลล์มาก ดังที่ทราบกันดีว่าสิ่งมีชีวิตในเซลล์มีจีโนม DNA แบบเกลียวคู่เท่านั้น - เชิงเส้นหรือแบบวงกลม และจีโนมของไวรัสสามารถแสดงได้ด้วย RNA หรือโมเลกุล DNA ทั้งแบบสายเดี่ยวและคู่แบบเส้นตรงหรือแบบวงกลม นอกจากนี้ยังมีระบบที่ใช้การถอดรหัสแบบย้อนกลับอีกด้วย ดังนั้นในรีโทรไวรัส (เช่น ออนโคไวรัสบางชนิด, เอชไอวี) และพารารีโทรไวรัส (ไวรัสตับอักเสบบี, โมเสกกะหล่ำดอก ฯลฯ) หนึ่งในสายโซ่ DNA ของจีโนมจะถูกสังเคราะห์บนเทมเพลต RNA ไวรัสต่างจากสิ่งมีชีวิตในเซลล์ที่ใช้วิธีการจัดเก็บและแสดงข้อมูลทางพันธุกรรมที่เป็นไปได้ตามทฤษฎีทั้งหมด
ข้อโต้แย้งที่สำคัญประการที่สองในการพิจารณาว่าไวรัสมีต้นกำเนิดมาจากเซลล์ก็คือ มียีนของไวรัสจำนวนมากที่ไม่มีอยู่ในสิ่งมีชีวิตในเซลล์ สิ่งมีชีวิตระดับเซลล์ไม่เพียงแต่วิวัฒนาการมาจากไวรัสเท่านั้น แต่ยังสืบทอด (และสืบทอดต่อ) ส่วนสำคัญของสารพันธุกรรมจากไวรัสอีกด้วย สิ่งที่น่าสนใจเป็นพิเศษในเรื่องนี้คือไวรัสภายนอก (ส่วนหนึ่งของจีโนมของ RNA หรือไวรัส DNA ที่รวมอยู่ในจีโนมของเซลล์) ซึ่งยีนที่ได้มาจากรีโทรไวรัสมีอิทธิพลเหนือกว่า เชื่อกันว่าสัตว์เลี้ยงลูกด้วยนมสืบทอดจีโนมมากกว่าครึ่งหนึ่งจากไวรัสและญาติที่ใกล้ที่สุด - องค์ประกอบทางพันธุกรรมที่ "เห็นแก่ตัว" เช่น พลาสมิดและทรานสโพซัน ดังนั้นไวรัสจึงเป็นพ่อแม่ร่วมของมนุษย์ บ่อยครั้งที่ลำดับยีนของไวรัสภายนอกซึ่งพบได้ในจีโนมมนุษย์จำนวนมาก มีการเปลี่ยนแปลงและไม่มีรหัสสำหรับโปรตีนอีกต่อไป มีเหตุผลที่ดีที่จะเชื่อได้ว่าลำดับดังกล่าวเกี่ยวข้องกับการควบคุมยีนของเซลล์ แม้ว่าการทำงานทางชีววิทยาจำเพาะของยีนนั้นมักจะไม่ทราบแน่ชัดก็ตาม อย่างไรก็ตาม เรารู้บางสิ่งที่สำคัญ ตัวอย่างเช่น โปรตีนซินซิตินซึ่งถูกเข้ารหัสโดยยีนซองจดหมายของรีโทรไวรัสภายนอกตัวหนึ่ง จำเป็นสำหรับการหลอมรวมของเซลล์ในระหว่างการก่อตัวของรก ซึ่งหมายความว่าทั้งมนุษย์และสัตว์ในรกไม่สามารถเกิดมาได้หากไม่มีไวรัสภายในร่างกายนี้ มีอีกตัวอย่างที่สำคัญคือ ปรากฎว่าส่วนประกอบของจีโนมของไวรัสภายนอกตัวหนึ่งควบคุมการแสดงออกของโพรลีนดีไฮโดรจีเนสในบางภูมิภาคของระบบประสาทส่วนกลาง เอนไซม์นี้อาจมีบทบาทสำคัญในวิวัฒนาการของสมองมนุษย์ หากการแสดงออกของเอนไซม์นี้หยุดชะงักอันเป็นผลมาจากการกลายพันธุ์ จะเกิดอาการป่วยทางจิต รวมถึงโรคจิตเภทด้วย ไวรัสและญาติของพวกมันยังมีบทบาทสำคัญในการถ่ายโอนยีนของเซลล์ในแนวนอนจากสิ่งมีชีวิตหนึ่งไปยังอีกสิ่งมีชีวิตหนึ่ง
อย่างไรก็ตาม แม้ว่าไวรัสจะมีบทบาทสำคัญในวิวัฒนาการ แต่ไวรัสยังเป็นที่รู้จักกันดีว่าเป็นเชื้อโรคของมนุษย์ สัตว์ และพืช (อย่างไรก็ตาม นี่คือสาเหตุว่าทำไมพวกมันถึงถูกค้นพบครั้งแรก) แล้วเราจะพูดถึงธรรมชาติของการเกิดโรคจากไวรัส ไวรัส (โดยเฉพาะไวรัสยูคาริโอต) ไม่มี "ความปรารถนา" พิเศษที่จะทำร้ายโฮสต์ และฆ่ามันได้น้อยมาก และในหลายกรณี ไวรัสอยู่ร่วมกันอย่างสันติและเป็นมิตรกับเซลล์ เหตุใดไวรัสจำนวนมากจึงเป็นอันตราย? คำอธิบายตามปกติคือพยาธิสภาพของเซลล์ที่ติดเชื้อเกิดจากการ "ปล้น" ทรัพยากรของเซลล์ (วัสดุและโครงสร้าง) ซึ่งไวรัสมุ่งตรงไปที่ความต้องการในการสืบพันธุ์ของมันเอง อย่างไรก็ตาม อันตรายที่ยิ่งใหญ่ที่สุดสามารถเกิดขึ้นได้จากการดำเนินการป้องกันที่ไม่เหมาะสมของโฮสต์และกิจกรรมการป้องกันของไวรัส ซึ่งไม่เกี่ยวข้องโดยตรงกับการแพร่พันธุ์
กลไกการป้องกันและการตอบโต้
กลไกการป้องกันหลักของเซลล์ที่ติดเชื้อคืออะไร? สิ่งเหล่านี้เป็นองค์ประกอบของภูมิคุ้มกันโดยกำเนิด: การเสื่อมของ RNA (ไวรัสและเซลล์) การยับยั้งการสังเคราะห์โปรตีน (ทั้งไวรัสและเซลล์) การทำลายตนเอง (การตายของเซลล์และการเสียชีวิตตามโปรแกรมประเภทอื่น) และสุดท้ายคือการอักเสบ จริงๆ แล้ว ไวรัสจำนวนมากค้นพบการมีอยู่ของพวกมันในลักษณะนี้ - เนื่องมาจากการอักเสบที่เกิดขึ้น (โรคไข้สมองอักเสบ โรคปอดบวม ฯลฯ) เซลล์ต่อสู้กับไวรัสโดยขัดขวางกระบวนการเผาผลาญและ/หรือโครงสร้างของมันเอง และกลไกการป้องกันของมันมักจะสร้างความเสียหายในตัวเอง คุณอาจพูดได้ว่าคนที่เสียชีวิตด้วยโรคโปลิโอ (และน้อยกว่า 1% เสียชีวิต) ฆ่าตัวตายขณะต่อสู้กับการติดเชื้อ
ในการตอบสนองต่อการป้องกันระดับเซลล์ ไวรัสจะพัฒนาเพื่อพัฒนาการป้องกัน และมีการแข่งขันทางอาวุธระหว่างไวรัสกับเซลล์ ยาเหล่านี้มุ่งเป้าไปที่กระบวนการเผาผลาญทั่วไปที่เป็นรากฐานของปฏิกิริยาการป้องกันของเซลล์เป็นหลัก สิ่งนี้คือการยับยั้งการสังเคราะห์ RNA และโปรตีนของเซลล์ การหยุดชะงักของโครงสร้างพื้นฐานภายในเซลล์และการขนส่งเซลล์ การปราบปรามหรือในทางกลับกัน การเปิดตัวของการตายของเซลล์และกลไกอื่น ๆ ที่ทำให้เซลล์ตายตามโปรแกรม ดังนั้นกลยุทธ์การป้องกันของไวรัสจึงคล้ายคลึงกับพฤติกรรมการป้องกันของเซลล์ในหลาย ๆ ด้าน นักมวยปล้ำใช้เทคนิคเดียวกันและบรรลุเป้าหมายเดียวกัน ตัวอย่างเช่น เซลล์ซึ่งยับยั้งการสังเคราะห์โปรตีนของไวรัส ใช้อินเตอร์เฟอรอน และเพื่อยับยั้งการก่อตัวของเซลล์ ในทางกลับกัน ไวรัสก็จะยับยั้งการสังเคราะห์โปรตีนในเซลล์ ขึ้นอยู่กับสถานการณ์ด้านใดด้านหนึ่งหรือผลประโยชน์อื่น ๆ ปรากฎว่าส่วนสนับสนุนหลักของพยาธิวิทยาไม่ใช่การแพร่พันธุ์ของไวรัสเช่นนี้ แต่เป็นการเผชิญหน้าระหว่างการป้องกันเซลล์และการป้องกันไวรัส ในพยาธิวิทยาของพืช แนวคิดเรื่อง "ความอดทน" มีมานานแล้ว: ไวรัสที่ทำให้เกิดโรคสามารถแพร่กระจายในพืชที่ติดเชื้อได้อย่างแข็งขันโดยไม่ก่อให้เกิดอาการเจ็บปวด
ด้านล่างนี้เราจะพูดถึงไวรัส RNA เป็นหลัก (นี่คือตัวอย่างที่ง่ายกว่า) เมื่อไวรัส RNA เข้าสู่เซลล์จะเปิดเผยการมีอยู่ของมันได้อย่างไร? และเซลล์จะรู้ได้อย่างไรว่ามีไวรัสเข้ามา? คุณสมบัติหลักเนื่องจากการที่เซลล์ "เข้าใจ" นี่คือ RNA แบบเกลียวคู่ของไวรัสซึ่งโดยหลักการแล้วสามารถก่อตัวได้ในเซลล์ที่ไม่ติดเชื้อ แต่ไม่ใช่ในปริมาณและตำแหน่งดังกล่าว ในบางกรณี เซลล์ยังจดจำ RNA สายเดี่ยวของไวรัส และบางครั้ง (บ่อยน้อยกว่ามาก) ของโปรตีนของไวรัส สิ่งสำคัญคือการรับรู้ RNA ของไวรัสนั้นไม่เฉพาะเจาะจง: การมี RNA แบบเกลียวคู่ "รับรู้" เซลล์อาจ "คิด" ว่ามีไวรัสเข้ามา แต่ไม่รู้ว่าอันไหน RNA ถูกจับโดยเซ็นเซอร์สองประเภท: คล้ายค่าผ่านทาง (จากภาษาอังกฤษ. เหมือนเก็บเงินและจากเขา ค่าผ่านทาง- โดดเด่น) ตัวรับและ RNA helicase เฉพาะทาง เกี่ยวข้องกับกลไกการป้องกันจำนวนหนึ่งในระดับการถอดเสียง รวมถึงการก่อตัวของอินเตอร์เฟอรอน นอกจากนี้ RNA ของไวรัสยังได้รับการยอมรับโดย "ผู้ดำเนินการ" - PKR โปรตีนไคเนสที่ขึ้นกับ RNA แบบสองเกลียวซึ่งฟอสโฟรีเลชั่นปัจจัยการเริ่มต้นการแปลบางส่วนจึงยับยั้งการสังเคราะห์โปรตีน oligoadenylate synthetase (OAS) ซึ่งกระตุ้น RNase L ซึ่งแยก RNA; ระบบการรบกวน RNA นำไปสู่การย่อยสลาย RNA และการหยุดชะงักของการแปล
เนื่องจากไวรัสได้รับการยอมรับว่าเป็นสิ่งที่ไม่เฉพาะเจาะจง เซลล์จึงไม่สามารถทราบ "ความตั้งใจ" ของมันได้ และโดยทั่วไปแล้ว มันคงเป็นไปไม่ได้เลยที่จะมีระบบการป้องกันโดยธรรมชาติสำหรับไวรัสใดๆ ก็ตามที่เป็นไปได้ ซึ่งหมายความว่าเซลล์สามารถต่อสู้กับไวรัสได้ด้วยเทคนิคมาตรฐานเท่านั้น ดังนั้น การดำเนินการป้องกันจึงมักจะไม่สมส่วนกับภัยคุกคามที่มีอยู่ อย่างไรก็ตาม หากปฏิกิริยาการป้องกันของเซลล์ไม่จำเพาะเจาะจง ทำไมไวรัสต่างชนิดกันจึงทำให้เกิดโรคต่างกัน ประการแรก ไวรัสแต่ละตัวสามารถแพร่เชื้อได้เฉพาะเซลล์บางประเภทในสิ่งมีชีวิตที่เฉพาะเจาะจงเท่านั้น นี่เป็นเพราะความจริงที่ว่าเพื่อที่จะเจาะเซลล์นั้นจะต้องมีปฏิกิริยากับตัวรับเซลล์ที่ "เหมาะสม" สำหรับมัน นอกจากนี้ สำหรับการแพร่พันธุ์ของไวรัส จำเป็นต้องมีสภาพแวดล้อมภายในเซลล์บางอย่าง (มักจำเป็นต้องใช้โปรตีนของเซลล์จำเพาะ) ประการที่สอง แม้ว่าปฏิกิริยาการป้องกันของเซลล์จะเป็นมาตรฐาน แต่การป้องกันของไวรัสนั้นส่วนใหญ่ขึ้นอยู่กับแต่ละบุคคล แม้ว่าจะมุ่งเป้าไปที่กลไกมาตรฐานของเซลล์ก็ตาม
ในพืช การรบกวน RNA มีบทบาทสำคัญมากในฐานะกลไกการต้านไวรัส RNA แบบเกลียวคู่เกิดขึ้นจาก RNA ของไวรัส (ปัจจัยสำคัญที่เซลล์เรียนรู้เกี่ยวกับการมีอยู่ของไวรัส) ด้วยการมีส่วนร่วมของส่วนประกอบของระบบการรบกวน RNA - เอนไซม์ Dicer ซึ่งตัด RNA แบบเกลียวคู่นี้ออกเป็นชิ้นส่วนที่มีความยาวนิวคลีโอไทด์ 21-25 คู่และจากนั้น RISC ที่ซับซ้อนของ RNA- โปรตีน - ในที่สุดชิ้นส่วน RNA สั้นแบบเกลียวเดี่ยวก็คือ เกิดขึ้น การผสมพันธุ์กับ RNA ของไวรัส ทำให้เกิดความเสื่อมหรือยับยั้งการแปลความหมาย กลไกการป้องกันนี้มีประสิทธิภาพแต่สามารถทำลายเซลล์ได้ ดังที่เห็นได้ชัดเจนในตัวอย่างของไวรอยด์ สิ่งเหล่านี้คือเชื้อโรคพืช ซึ่งเป็นโมเลกุล RNA แบบเกลียวเดี่ยวแบบวงกลมขนาดสั้น (หลายร้อยนิวคลีโอไทด์) ที่ไม่มีเปลือกโปรตีนปกคลุมอยู่ ไวรอยด์ไม่ได้เข้ารหัสโปรตีน แต่อาจทำให้เกิดอาการรุนแรงในพืชที่ติดเชื้อได้ สิ่งนี้เกิดขึ้นเพราะเซลล์กำลังปกป้องตัวเอง RNA แบบเกลียวคู่ของไวรอยด์ที่เกิดขึ้นจะถูกเปิดเผยต่อส่วนประกอบทั้งหมดของระบบรบกวน RNA ส่งผลให้เกิดการก่อตัวของชิ้นส่วน RNA แบบเกลียวเดี่ยวที่ไม่ผสมกับ RNA ของไวรัส แต่กับ RNA ของเซลล์ สิ่งนี้นำไปสู่การย่อยสลายและการพัฒนาอาการของโรค อย่างไรก็ตาม ไวรัสพืชหลายชนิดเข้ารหัสโปรตีนหลายชนิดที่รบกวนการรบกวน RNA ( ตัวยับยั้งไวรัสของการเงียบ RNA - VSR). พวกมันยับยั้งการรับรู้และความแตกแยกของ RNA ของไวรัส หรือระงับการก่อตัวและการทำงานของ RISC complex ดังนั้นโปรตีน VSR เหล่านี้สามารถรบกวนกลไกการรบกวน RNA ที่สำคัญทางสรีรวิทยา (ไม่ใช่ไวรัส) ทำให้เกิดอาการทางพยาธิวิทยา
โปรตีนความปลอดภัย
การป้องกันไวรัส โดยเฉพาะพิคอร์นาไวรัส ซึ่งเป็นเชื้อโรคที่มี RNA ขนาดเล็ก ส่วนใหญ่ขึ้นอยู่กับการทำงานของโปรตีนดังกล่าว โดยเฉพาะอย่างยิ่งกลุ่มใหญ่นี้รวมถึงเชื้อโรคของโปลิโอไมเอลิติส, โรคตับอักเสบเอ, โรคปากและเท้าเปื่อย ฯลฯ ลักษณะเฉพาะของไวรัสเหล่านี้คือโปรตีนของพวกมันถูกสังเคราะห์ในรูปแบบของโพลีโปรตีนเดี่ยวจากข้อยกเว้นที่หายาก จากนั้นจึงสร้างโปรตีนที่เจริญเต็มที่ขึ้นมา ในหมู่พวกเขาสามารถแยกแยะได้สามกลุ่ม โปรตีนชนิดแรกประกอบด้วยโปรตีนสำคัญซึ่งมีความสำคัญพร้อมหน้าที่คงที่ซึ่งรับประกันการแพร่พันธุ์ของไวรัสโดยตรง: RNA polymerases ที่ขึ้นกับ RNA ซึ่งจำเป็นสำหรับการจำลองแบบของจีโนมของไวรัส โปรตีนแคปซิดที่สร้างเปลือกโปรตีนของไวรัส โปรตีเอสที่เกี่ยวข้องในกระบวนการเปลี่ยนโพลีโปรตีนเป็นโปรตีนที่โตเต็มที่ โปรตีนวีพีจี ( จีโนมโปรตีนของไวรัสเชื่อมโยงกัน- โปรตีนของไวรัสที่เชื่อมต่อกับจีโนม) ซึ่งทำหน้าที่เป็นไพรเมอร์สำหรับการสังเคราะห์โมเลกุล RNA Helicase เป็นเอนไซม์ที่มีคุณค่ามากซึ่งมีอยู่ใน picornaviruses แต่มีบทบาทที่ไม่ชัดเจนนัก กลุ่มที่สองยังรวมถึงโปรตีนที่สำคัญด้วย แต่กลุ่มที่ทำงาน "เสริม" - โปรตีน "นำทาง" ที่ไม่ชอบน้ำ 2B และ 3A พวกมันนำโปรตีนสำคัญไปยังจุดหมายปลายทางและช่วยสร้างสภาพแวดล้อมภายในเซลล์ที่เหมาะสมที่สุดสำหรับการสืบพันธุ์ของไวรัส กลุ่มที่สาม ได้แก่ โปรตีนผู้นำ L ซึ่งค้นพบในห้องปฏิบัติการของเราเมื่อ 30 ปีที่แล้ว และโปรตีน 2A เราเรียกพวกมันว่าโปรตีน "ความปลอดภัย" ( ความปลอดภัย- ความปลอดภัย). นี่คือ "อาวุธ" ต่อต้านการป้องกันเฉพาะของ picornaviruses โดยทั่วไปแล้ว โปรตีนทั้งสามประเภทนี้สามารถต่อสู้กับกลไกการป้องกันของเซลล์ได้ แต่โปรตีนหลักและโปรตีนนำทางทำงานนี้นอกเวลาเพราะมีหน้าที่สำคัญอื่นๆ ที่โครงสร้างและหน้าที่ของมันต้องรองรับ ด้วยเหตุนี้ ความสามารถในการป้องกันจึงถูกจำกัดโดยความจำเป็นในการทำงานขั้นพื้นฐาน แต่กระรอก "ความปลอดภัย" ทำงานเต็มเวลาตามความสามารถพิเศษ - วิวัฒนาการ "จ้าง" พวกมันเพื่อ "ความปลอดภัย" โดยเฉพาะ (ต่อมาบางคน "เรียนรู้" ที่จะทำอย่างอื่น) พวกเขาสามารถมีโครงสร้างที่จำเป็นในการปฏิบัติหน้าที่ได้
หน้าที่ที่สำคัญที่สุดอย่างหนึ่งของโปรตีนรักษาความปลอดภัยคือพวกมันมีส่วนร่วมในการกำหนดชะตากรรมของเซลล์ที่ติดเชื้อ มีหลายทางเลือกสำหรับการตายของมัน แต่กลไกหลักสองอย่างที่รู้จักกันดีที่สุดคือเนื้อร้ายและการตายของเซลล์ ซึ่งมีลักษณะทางสัณฐานวิทยาและชีวเคมีแตกต่างกัน ในระหว่างการตายของเซลล์ เซลล์จะถูก lysed และเนื้อหาจะถูกเทลงในช่องว่างระหว่างเซลล์ ในระหว่างการตายของเซลล์ ส่วนที่ยื่นออกมาที่มองเห็นได้ชัดเจนจะเกิดขึ้นบนพื้นผิวของมัน DNA ของมันจะถูกสลายไปเป็นชิ้นส่วนของนิวคลีโอโซม และท้ายที่สุด เซลล์ก็จะถูกแยกส่วนออกเป็นร่างกายอะพอพโทซิสแต่ละตัวซึ่งจำกัดอยู่ที่พลาสมาเมมเบรน มันสำคัญมากว่าเซลล์จะตายอย่างไร เมื่อเนื้อตายจะเกิดการอักเสบในการป้องกัน แต่ไวรัสจะออกจากเซลล์และแพร่กระจาย การตายของเซลล์จะทำให้การแพร่กระจายของไวรัสมีจำกัด และมักไม่มีปฏิกิริยาการอักเสบ ตามกฎแล้วการตายของเซลล์ที่ติดเชื้อถือเป็นการเสียสละตนเองซึ่งจำกัดการแพร่พันธุ์ของไวรัส
เราพบว่าการติดเชื้อ picornaviruses โดยเฉพาะโปลิโอไวรัส (โปลิโอไวรัส) กระตุ้นให้เกิดโปรแกรมการตายของเซลล์ สิ่งนี้เกิดขึ้นตามวิถีคลาสสิกสายหนึ่ง เมื่อไซโตโครมถูกปล่อยออกจากไมโตคอนเดรีย คและมีการเปิดใช้งานน้ำตกของเอนไซม์โปรตีโอไลติก แคสเปส แต่ในทางกลับกัน ปรากฎว่าไวรัสมีกลไกต่อต้านการตายของเซลล์ นั่นคือความสามารถในการยับยั้งปฏิกิริยาอะพอพโทติกของเซลล์ ดังนั้นเซลล์ HeLa ที่ติดเชื้อไวรัสโปลิโอหรือไวรัสไข้สมองอักเสบ (เช่น picornavirus) จะตายโดยมีสัญญาณของเนื้อร้าย แต่ถ้าคุณปิด "อาวุธ" ต่อต้านการตายของเซลล์ (ระงับการสังเคราะห์โปรตีนของไวรัส) เซลล์จะตายจากการตายของเซลล์ (การเสียสละตนเอง) ในไวรัสทั้งสองชนิด โปรตีน "ความปลอดภัย" ทำหน้าที่เป็นอาวุธดังกล่าว อย่างไรก็ตาม ในไวรัสไข้สมองอักเสบ บทบาทนี้เล่นโดยโปรตีน L และในไวรัสโปลิโอคือโปรตีน 2A โปรตีนผู้นำไม่มีการทำงานของเอนไซม์ ในขณะที่โปรตีน 2A เป็นโปรตีเอส พวกมันไม่มีอะไรที่เหมือนกันทั้งเชิงโครงสร้างหรือทางชีวเคมี แต่ทั้งสองมีฤทธิ์ต้านอะพอพโทติคตามกลไกระดับโมเลกุลที่แตกต่างกัน

กลไกการป้องกันการป้องกันอีกประการหนึ่งของโปรตีนความปลอดภัยของ picornavirus คือการหยุดชะงักของการขนส่งนิวเคลียส - ไซโตพลาสซึม [10–12] เราได้แสดงให้เห็นว่าเมื่อติดเชื้อไวรัสเหล่านี้ การซึมผ่านของเยื่อหุ้มนิวเคลียสจะเพิ่มขึ้น และการแลกเปลี่ยนโมเลกุลขนาดใหญ่ระหว่างไซโตพลาสซึมและนิวเคลียสจะหยุดชะงัก และหากโครงสร้างเซลล์เสียหาย ก็จะไม่สามารถเปิดกลไกควบคุมเพื่อต่อสู้กับไวรัสได้ ในโปลิโอไวรัส โปรตีนความปลอดภัย 2A ขัดขวางการขนส่งนิวเคลียส-ไซโตพลาสซึมโดยการไฮโดรไลซ์นิวคลีโอปอริน ซึ่งเป็นส่วนประกอบของรูพรุนนิวเคลียร์ และโปรตีนผู้นำของไวรัสไข้สมองอักเสบทำงานได้ - มันส่งผลต่อน้ำตกของเซลล์ของฟอสโฟรีเลชั่นของนิวคลีโอปอริน [,]
หน้าที่ต่อต้านการป้องกันของโปรตีน "ความปลอดภัย" สามารถแสดงออกได้ด้วยวิธีอื่น ดังนั้น L-proteins ของ cardioviruses (รวมถึงไวรัส encephalomyocarditis) และ 2A-proteins ของ enteroviruses (รวมถึงโปลิโอไวรัส) ยับยั้งการก่อตัวของ interferon และออกฤทธิ์ถูกยับยั้งโดย L-protein ของไวรัสโรคปากและเท้าเปื่อยและโปรตีน 2A ของไวรัสโปลิโอ อย่างไรก็ตาม โปรตีน "ความปลอดภัย" ของ picornaviruses นั้นไม่สำคัญ ผู้พิทักษ์ทั้งสองสามารถถูกลบออกได้หรืออาจทำให้เกิดการลบจำนวนมากได้ (เช่นในกรณีของโปรตีน L ของคาร์ดิโอไวรัส, โปรตีน 2A ของไวรัสตับอักเสบเอ และคาร์ดิโอไวรัส) โดยที่ไวรัสไม่สามารถทำงานได้
การปลดอาวุธซึ่งกันและกัน

อะไรคือผลที่ตามมาจากการหยุดใช้งานโปรตีน "ความปลอดภัย" ของไวรัสสำหรับเซลล์? ในด้านหนึ่งความไวของไวรัสต่อกลไกการป้องกันภูมิคุ้มกันของเซลล์โดยธรรมชาติจะเพิ่มขึ้น แต่ในทางกลับกัน กิจกรรมการทำร้ายตัวเองและฆ่าตัวตายก็จะเพิ่มขึ้นเช่นกัน จะเกิดอะไรขึ้นหากคุณปิดกลไกการป้องกันของเซลล์และไวรัสไปพร้อม ๆ กัน? เราศึกษาสถานการณ์นี้โดยใช้ตัวอย่างปฏิสัมพันธ์ระหว่าง mengovirus (สายพันธุ์ของไวรัสไข้สมองอักเสบ) และเซลล์ HeLa เมื่อติดไวรัสชนิดป่า พวกมันจะตายจากเนื้อร้ายอย่างรวดเร็ว และหากไวรัสถูกปลดอาวุธบางส่วน (โปรตีนผู้นำถูกปิดใช้งาน) เซลล์ของ HeLa จะมีชีวิตยืนยาวขึ้นเล็กน้อยและไม่ได้ตายจากเนื้อร้าย แต่จากการตายของเซลล์ เมื่อการป้องกันซึ่งกันและกันลดลง (การตายของเซลล์ในเซลล์ถูกปิดโดยสารประกอบทางเคมีที่ยับยั้งแคสเปส และโปรตีนผู้นำของไวรัสถูกปิดใช้งาน) แม้หลังจากผ่านไปเป็นระยะเวลานานเป็นสองเท่า เซลล์จะรู้สึกดีขึ้นอย่างมีนัยสำคัญมากกว่าเซลล์ที่ไม่ได้เป็นเช่นนั้น ปลดอาวุธ และการแพร่พันธุ์ของไวรัส (ทั้งการเปลี่ยนแปลงและการเก็บเกี่ยว) ดำเนินไปในลักษณะเดียวกันทุกประการ โดยไม่คำนึงว่าเพียงการป้องกันเท่านั้นที่ถูกปิด หรือการป้องกันระดับเซลล์จะถูกลบออกในเวลาเดียวกันหรือไม่ ปรากฎว่าในเซลล์ที่ยังไม่มีความเสียหายทางพยาธิวิทยาร้ายแรง (ที่เรียกว่าผลทางไซโตพาธี) อนุภาคไวรัสจำนวนมากสามารถก่อตัวได้แล้ว ดังนั้นจึงไม่จำเป็นต้องทำลายเซลล์เพื่อให้ไวรัสสามารถแพร่พันธุ์ได้ ดังนั้นกลยุทธ์ที่มีประสิทธิภาพสำหรับการรักษาด้วยยาต้านไวรัสที่มุ่งบรรเทาอาการของโรคอาจเป็นการปราบปรามการป้องกันทั้งไวรัสและเซลล์พร้อมกัน
โปรแกรมความตาย
การทดลองชุดนี้ของเรายังให้โอกาสในการเจาะลึกลงไปในธรรมชาติของการตายแบบตายที่เกิดจากไวรัส มันคืออะไร - การฆ่าเซลล์ด้วยไวรัสหรือการฆ่าตัวตาย (การเสียสละตนเอง) เมื่อมันตัดสินใจว่าเพื่อประโยชน์ส่วนรวมมันสมควรที่จะตายมากกว่า? ต่อไปนี้อาจมีรอยโรคตาย:
- พลาสมาเมมเบรน (การซึมผ่านเพิ่มขึ้น, รูปแบบ "แผลพุพอง")
- ไซโตพลาสซึม (การเปลี่ยนแปลงของไมโครทูบูลและไมโครฟิลาเมนต์)
- นิวเคลียส (หดตัว, ผิดรูป, โครมาตินควบแน่น),
- กิจกรรมการเผาผลาญ (ปฏิกิริยาการลดลงขึ้นอยู่กับ NADH การเปลี่ยนแปลงความสามารถในการมีชีวิต)
เมื่อปิดการตายของเซลล์ (การเติมตัวยับยั้งแคสเปสทางเคมี) การเปลี่ยนแปลงของเนื้อร้ายต่างๆ ขึ้นอยู่กับว่าโปรตีนผู้นำของไวรัสทำงานหรือไม่ ตัวอย่างเช่น หากปิดใช้งาน ความสามารถในการซึมผ่านของเยื่อหุ้มเซลล์จะไม่เปลี่ยนแปลง "แผลพุพอง" ที่เกี่ยวข้องกับความไม่สมดุลในสมดุลออสโมติกจะไม่ปรากฏขึ้น และไม่มีรอยโรคเนื้อตายอื่นๆ อีกจำนวนหนึ่งเกิดขึ้น คำอธิบายหนึ่งที่เป็นไปได้สำหรับผลกระทบนี้คือ แอลโปรตีนออกฤทธิ์กับหลายเป้าหมายในช่องเซลล์ต่างๆ แต่เนื่องจากโปรตีนมีขนาดเล็กและไม่มีกิจกรรมของเอนไซม์ จึงมีแนวโน้มว่าเป้าหมายโดยตรงจะมีขนาดเล็กกว่ามาก เราตั้งสมมติฐานว่าโปรตีนผู้นำออกฤทธิ์กับองค์ประกอบเซลล์สำคัญหนึ่งองค์ประกอบหรือมากกว่านั้นที่ควบคุมชะตากรรมของเซลล์ และด้วยเหตุนี้ โปรแกรมการตายของเซลล์จึงเริ่มต้นขึ้น ซึ่งมีหน้าที่รับผิดชอบต่อการเปลี่ยนแปลงทางพยาธิวิทยาที่ระบุไว้ส่วนใหญ่ ดังนั้นจึงไม่ใช่ไวรัสที่ฆ่าเซลล์แบบตายตัว แต่เซลล์เองก็ฆ่าตัวตาย (เป็นการเสียสละตนเอง) มุมมองนี้สอดคล้องกับแนวคิดใหม่ ซึ่งนอกเหนือจากการตายของเซลล์แล้ว ยังมีประเภทการตายของเซลล์ที่ตั้งโปรแกรมไว้ (เข้ารหัสในจีโนมของเซลล์) ที่มีความสำคัญทางสรีรวิทยาอีกหลายประเภท รวมถึงการตายของเซลล์ซึ่งคล้ายกับการตายของเซลล์
ดังนั้นการเสียสละตัวเองของเซลล์ในระหว่างการติดเชื้อไวรัสสามารถแสดงออกได้ว่าเป็นการตายของเซลล์หรือการตายของเซลล์ การตายของเซลล์อาจเป็นปฏิกิริยาป้องกันของเซลล์ต่อการติดเชื้อไวรัส และไม่ใช่เฉพาะในกรณีของพิคอร์นาไวรัสเท่านั้น กลไกใดที่เป็นประโยชน์ต่อไวรัสมากกว่านั้นขึ้นอยู่กับสภาวะ เราเห็นว่าผลในการต่อต้านการป้องกันสามารถแสดงออกมาในรูปแบบของ "การเปลี่ยนเส้นทาง" ของกลไกที่เข้ารหัสในจีโนมของเซลล์ นี่เป็นวิธีการป้องกันที่สำคัญ (แม้ว่าจะไม่ใช่วิธีเดียว) และเป็นหนึ่งในกลไกหลักของการทำให้เกิดโรคของไวรัส โปรแกรมการตายของเซลล์และเนื้อร้ายที่เกิดจากไวรัสแข่งขันกัน เราได้แสดงให้เห็นว่าเมื่อเซลล์ HeLa ติดเชื้อไวรัสโปลิโอ การตายของเซลล์จะถูกกระตุ้นในครั้งแรก จากนั้นจะถูกระงับและวิถีทางของการตายของเซลล์จะถูกเปิดขึ้น ดังนั้นการติดเชื้อของเซลล์ด้วยไวรัสจะกระตุ้นการดำเนินการป้องกันหลายอย่างซึ่งมีกลไกการฆ่าตัวตายสองประการของการเสียชีวิตตามโปรแกรม - apoptotic และ necrotic จากนั้นการแข่งขันก็เกิดขึ้นระหว่างเส้นทางเหล่านี้: การปราบปรามหนึ่งในนั้นจะเป็นการเปิดใช้งานอีกเส้นทางหนึ่ง และในทางกลับกัน และทั้งหมดนี้ควบคุมโดยโปรตีนในเซลล์ โปรตีนของไวรัส (โดยหลักคือโปรตีน "ความปลอดภัย") รวมถึงปัจจัยภายนอก
การแข่งขันด้านอาวุธ
เนื่องจากเซลล์มีกลไกการป้องกัน และไวรัสก็มีกลไกการป้องกัน จึงมีการแข่งขันทางอาวุธระหว่างเซลล์เหล่านั้น ธรรมชาติที่ไม่อนุรักษ์นิยมของโปรตีนความปลอดภัยแสดงให้เห็นว่าพวกมันถูกดัดแปลงเพื่อต่อต้านกลไกการป้องกันของโฮสต์เฉพาะ ดังนั้นการเปลี่ยนแปลงอาจมาพร้อมกับการสูญเสียการทำงานของโปรตีน "ความปลอดภัย" และผลที่ตามมาคือการเพิ่มขึ้นของปฏิกิริยาการป้องกันของเจ้าบ้าน สิ่งนี้สามารถอธิบายการเกิดโรคพิเศษของ "ใหม่" ( ที่เพิ่งเกิดขึ้นใหม่- อุบัติใหม่) ไวรัส ดังนั้นไวรัสไข้หวัดใหญ่จึงเป็นไวรัสในลำไส้ของนกป่าที่ก่อให้เกิดโรคต่ำและแทบไม่เป็นอันตราย เมื่อติดเชื้อในคน อาจเกิดไข้หวัดสเปน ไข้หวัดนก หรือไข้หวัดหมูได้ ไวรัสซาร์สค่อนข้างปลอดภัยสำหรับค้างคาว แต่ในมนุษย์ ไวรัสทำให้เกิดอาการทางเดินหายใจเฉียบพลันรุนแรง ร่วมกับอัตราการเสียชีวิตสูง ในที่สุด เอชไอวี (หรือบรรพบุรุษของมัน) แทบไม่เป็นอันตรายต่อลิง แต่ในมนุษย์ เชื้อเอชไอวีทำให้เกิดโรคเอดส์ เป็นสิ่งสำคัญมากที่ไวรัสเหล่านี้จะไม่พัฒนาปัจจัยการทำให้เกิดโรคใหม่เมื่อย้ายไปยังโฮสต์ใหม่ (เพียงเป็นผลมาจากการกลายพันธุ์หลายอย่างที่ทำให้แน่ใจได้ว่าสามารถแทรกซึมเข้าไปในเซลล์ได้ ทำให้พวกมันมีความสามารถในการแพร่เชื้อในมนุษย์) กลไกที่เป็นไปได้อีกประการหนึ่งสำหรับความไม่สมดุลระหว่างไวรัสกับโฮสต์และการเกิดขึ้นของเชื้อโรคใหม่อาจเป็นการเปลี่ยนแปลงในอาวุธป้องกันไวรัส เช่น การสูญเสียอันเก่าหรือการได้มาซึ่งโปรตีน "ความปลอดภัย" ใหม่
อย่างไรก็ตาม การวิวัฒนาการร่วมกันในระยะยาวของโฮสต์และไวรัสควรนำไปสู่การลดการเกิดโรคของไวรัสในระยะหลัง (การลดอาวุธที่เป็นประโยชน์ร่วมกัน) ตัวอย่างคลาสสิกคือไวรัส myxoma/fibroma ในช่วงกลางศตวรรษที่ 19 กระต่ายยุโรปถูกนำมาที่ออสเตรเลีย พวกมันขยายพันธุ์อย่างรวดเร็วและกลายเป็นภัยคุกคามร้ายแรงต่อการเกษตร 100 ปีต่อมา ไวรัส fibroma/myxoma ที่ทำให้เกิดโรค (จากตระกูล poxvirus ซึ่งรวมถึงไวรัสไข้ทรพิษ) เริ่มถูกนำมาใช้เพื่อควบคุมจำนวนประชากร กระต่ายต่างมีปฏิกิริยาต่อไวรัสนี้แตกต่างกัน ในกระต่ายบราซิล สามสัปดาห์หลังการติดเชื้อ จะทำให้เกิดเนื้องอกที่ไม่ร้ายแรง - เนื้องอกไฟโบรมา (ก้อนเนื้อเฉพาะที่บนผิวหนัง) แต่กระต่ายยุโรปที่ไวต่อไวรัสนี้จะเป็นโรคร้ายแรงถึงชีวิตภายใน 10 วันหลังการติดเชื้อ
ไวรัสดังกล่าวแพร่ระบาดในออสเตรเลียในออสเตรเลียในฤดูร้อน โดยกระต่ายที่ติดเชื้อมากกว่า 99% เสียชีวิตในเวลาไม่ถึงสองสัปดาห์ ไวรัสที่มีความรุนแรงน้อยกว่ามีแนวโน้มที่จะอยู่รอดได้ในช่วงฤดูหนาว และสิ่งนี้นำไปสู่การเลือกสายพันธุ์ที่อ่อนแอ (ลดทอน) และหลังจากนั้นประมาณ 10 ปี อัตราการตายของกระต่ายยุโรปจากไวรัสที่วิวัฒนาการก็ลดลงครึ่งหนึ่ง ในเวลาเดียวกัน มีการคัดเลือกกระต่ายต้านทาน: อัตราการตายของพวกมันจากไวรัสดั้งเดิมลดลงประมาณสี่เท่า ในเวลาเพียงทศวรรษ (ซึ่งเป็นช่วงเวลาเล็กน้อยในแง่วิวัฒนาการ) ความสัมพันธ์ระหว่างเชื้อโรคและโฮสต์ได้รับการปรับปรุงประมาณ 10 เท่า แน่นอนว่านี่เป็นโครงการที่ค่อนข้างเรียบง่าย เนื่องจากการแข่งขันทางอาวุธไม่ได้หยุดลง: เพื่อตอบสนองต่อความต้านทานที่เพิ่มขึ้นในกระต่าย ความรุนแรงของไวรัสก็อาจเพิ่มขึ้นเช่นกัน อย่างไรก็ตาม นี่เป็นตัวอย่างที่ชัดเจนของบทบาทของปฏิสัมพันธ์ของไวรัสและสิ่งมีชีวิตในเซลล์ในการวิวัฒนาการของทั้งสองอย่าง ไวรัสและเซลล์ “สอน” ซึ่งกันและกัน และ “ความรู้” ที่ได้มานั้นได้รับการสืบทอดมา ในปี 2013 ผู้สำเร็จการศึกษาสองคนจากภาควิชาไวรัสวิทยาของมหาวิทยาลัยแห่งรัฐมอสโก Evgeny Kunin และ Valeryan Dolya ตีพิมพ์บทความเกี่ยวกับมุมมองวิวัฒนาการแบบ "ไวโรเซนทริค" ตามที่ความต้านทานและความร่วมมือของไวรัสและสิ่งมีชีวิตในเซลล์เป็นปัจจัยหลักใน วิวัฒนาการของพวกเขา
เรื่องราวของฉันไม่ได้ทำให้หัวข้อหมดสิ้น: ยังมีความรู้อีกมากมายเกี่ยวกับธรรมชาติของการเกิดโรคของไวรัส สิ่งที่เรารู้ส่วนใหญ่ได้รับการเรียนรู้ในช่วงไม่กี่ปีที่ผ่านมา และมีเหตุผลทุกประการที่จะคาดหวังความประหลาดใจใหม่ๆ เราสามารถและควรตำหนิไวรัสสำหรับการเจ็บป่วยร้ายแรง และจำเป็นต้องต่อสู้กับมัน แต่เราควรจะขอบคุณไวรัสสำหรับการดำรงอยู่และความหลากหลายของธรรมชาติของสิ่งมีชีวิต รวมถึงการดำรงอยู่ของมนุษย์ด้วย
ผู้เขียนรู้สึกขอบคุณเพื่อนร่วมงานของเขาในความร่วมมือทางวิทยาศาสตร์ - เจ้าหน้าที่ของสถาบันโปลิโอไมเอลิติสและโรคไข้สมองอักเสบจากไวรัส M. P. Chumakov RAMS มหาวิทยาลัยแห่งรัฐมอสโก M.V. Lomonosov, สถาบันโปรตีน RAS (พุชชิโน, เขตมอสโก), มหาวิทยาลัยบาเซิล (สวิตเซอร์แลนด์), มหาวิทยาลัยวิสคอนซิน (สหรัฐอเมริกา), มหาวิทยาลัย Nijmegen รัดโบดา (เนเธอร์แลนด์)
บทความนี้มีพื้นฐานมาจากการบรรยายที่โรงเรียน "ชีววิทยาสมัยใหม่และเทคโนโลยีชีวภาพแห่งอนาคต" (Zvenigorod, 26 มกราคม - 1 กุมภาพันธ์ 2014)
. Belov G. A. , Lidsky P. V. , Mikitas O. V. และคณะ การซึมผ่านของซองจดหมายนิวเคลียร์เพิ่มขึ้นแบบสองทิศทางจากการติดเชื้อโปลิโอและการเปลี่ยนแปลงของรูขุมขนนิวเคลียร์ // เจ. วิโรล. 2004. V. 78. หน้า 10166–10177. ดอย:10.1128/JVI.78.18.10166-10177.2004. Lidsky P. V. , Hato S. , Bardina M. V. และคณะ ความผิดปกติของการจราจรนิวคลีโอไซโตพลาสซึมที่เกิดจากคาร์ดิโอไวรัส // เจ. วิโรล. 2549 โวลต์ 80 หน้า 2705–2717 ดอย:10.1128/JVI.80.6.2705-2717.2006
. Bardina M.V., Lidsky P.V., Sheval E.V. และคณะ การจัดเรียงรูพรุนเชิงซ้อนของนิวเคลียร์ที่เกิดจาก Mengovirus: การแย่งชิงเครื่องจักรฟอสโฟรีเลชั่นของเซลล์ // เจ. วิโรล. 2552 V. 83 หน้า 3150–3161. ดอย:10.1128/JVI.01456-08
. Mikitas O. V. , Ivin Y. Y. , Golyshev S. A. และคณะ การปราบปรามการบาดเจ็บที่เกิดจากไวรัส lytic RNA (mengovirus) และการแยกตัวจากการสืบพันธุ์ของไวรัสโดยการลดอาวุธของเซลล์/ไวรัส // เจ. วิโรล. 2012. V. 86. หน้า 5574–5583. ดอย:10.1128/JVI.07214-11
. Agol V. I. , Belov G. A. , Bienz K. และคณะ การแข่งขันโปรแกรมการตายในเซลล์ที่ติดเชื้อไวรัสโปลิโอ: สวิตช์ความมุ่งมั่นในช่วงกลางของวงจรการติดเชื้อ // เจ. วิโรล. 2000 V. 74. หน้า 5534–5541. ดอย:10.1128/JVI.74.12.5534-5541.2000
. Agol V. I. ผลกระทบทางไซโตพาธี: อาการที่ปรับด้วยไวรัสของภูมิคุ้มกันโดยธรรมชาติ? // เทรนด์ไมโครไบโอล 2012. V. 20. หน้า 570–576. ดอย:10.1016/j.tim.2012.09.003
. Koonin E. V. , Dolja V. V. มุมมองแบบ virocentric ต่อวิวัฒนาการของชีวิต // สกุลเงิน ความคิดเห็น. วิโรล. 2013. ฉบับที่ 3. หน้า 546–557. ดอย:10.1016/j.coviro.2013.06.008
 ธรรมชาติและที่มาของไวรัส
ธรรมชาติและที่มาของไวรัส นามบัตร QR - นามบัตรที่ใช้รหัส QR ออกแบบโดยใช้เว็บแอปพลิเคชัน
นามบัตร QR - นามบัตรที่ใช้รหัส QR ออกแบบโดยใช้เว็บแอปพลิเคชัน แผนทางเทคนิครูปหลายเหลี่ยมของสถานที่
แผนทางเทคนิครูปหลายเหลี่ยมของสถานที่